Gėlas vanduo ir jo gyventojai. Samanos – senovinės ir svarbios
tema: „Apatiniai ir aukštesni augalai: dumbliai, bryofitai ir paparčiai“
Žemesni ir aukštesni augalai
Augalai skirstomi į dvi grupes arba dvi subkaralystes: žemesniuosius ir aukštesniuosius. Žemesniaisiais vadinami tokie augalai, kurių vegetatyvinis kūnas neskirstomas į organus, o aukštesniaisiais – augalai su specializuotais vegetatyviniais organais, susidedančiais iš skirtingų audinių.
Augalų taksonomija
Suprasti biologinę įvairovę leidžia sisteminis mokslas. Yra augalų, gyvūnų, grybų, bakterijų taksonomija.
Tiesą sakant, įprastame gyvenime žmogus taip pat sistemina. Taigi, puodelis, stiklinė, cukraus dubuo jungiami į kategoriją „arbatos indai“, o lėkštė, šakutė, šaukštas – į kategoriją „vakarienės indai“. Abi kategorijos yra sujungtos į platesnę kategoriją „indai“. Kartu su baldais indus, pavyzdžiui, galima sugrupuoti į dar platesnę kategoriją „namų apyvokos reikmenys“ ir kt.
Kuo žemesnis sisteminės kategorijos reitingas, tuo didesnis jų skaičius. Žinoma tik 15-16 augalų skyrių, o rūšių apie 350 000. Vienai šeimai gali priklausyti 1000 genčių. Gentys žinomos 2000-3000 rūšių. Tačiau yra genčių, turinčių tik vieną rūšį, ir šeimų, turinčių vieną gentį.
Rūšis yra pagrindinis taksonomijos vienetas. Vardindami augalą, paprastai turime omenyje rūšį. Tos pačios rūšies augalai gali susilaukti palikuonių, skirtingos rūšys, kaip taisyklė, negali. Todėl gamtoje kartu gyvenančios rūšys nesikryžmina ir gerai skiriasi. Kiekviena rūšis būtinai priklauso visoms kitoms sisteminėms kategorijoms. Pvz.: drebulė priklauso tuopų genčiai, gluosnių šeimai, gluosnių eilei, dviskilčių klasei, žydinčių augalų skyriui.
Rūšies pavadinimas paprastai susideda iš dviejų žodžių, kurių pirmasis reiškia gentį, o antrasis – pačią rūšį (pavyzdžiui, karpinis beržas, kaustinis ranunculus, juodasis serbentas; išimtis yra drebulė, pomidoras, bulvė).
Mokslinėse knygose vardai būtinai rašomi lotyniškai (tai suprantama viso pasaulio mokslininkams).
Tos pačios rūšies kultūriniuose augaluose dažnai yra daug veislių. Pavyzdžiui, naminėje obelėje jų yra keli tūkstančiai. Įvairovė yra žmogaus veiklos rezultatas. Veislės, skirtingai nei rūšys, gali kryžmintis viena su kita.
Dumbliai: jų ekologija ir reikšmė
Bendrosios dumblių savybės. Dumbliai nuo kitų augalų organizmų skiriasi šiais būdais:
Dumbliai arba žemesni augalai
Jie daugiausia gyvena vandenyje.
Jie neturi vegetatyvinių organų.
Reprodukciniai organai yra vienaląsčiai.
Dažymas įvairus.
aukštesni augalai
Jie gyvena sausumoje (gyvenimas vandenyje yra antraeilis dalykas).
Jie turi vegetatyvinius organus.
Reprodukciniai organai yra daugialąsčiai.
Lapai visada žali.
Dumbliai gali būti vienaląsčiai ir daugialąsčiai, kartais dideli, iki kelių dešimčių metrų. Jie fotosintetina ir turi chlorofilo, bet dažnai ir kitų pigmentų. Dumblių spalva gali būti žalia, gelsva, ruda, raudona. Dumblių ląstelė turi tokias pačias dalis kaip ir aukštesniuose augaluose. Jų fotosintetiniai plastidai, nuo kurių priklauso spalva, vadinami chromatoforais. Dumbliai dauginasi vegetatyviniu, nelytiniu ir lytiniu būdu.
Žalieji dumbliai, gyvenantys gėlame vandenyje. Daugiausia žaliųjų dumblių gyvena gėlo vandens telkiniuose, tačiau jūrose jų palyginti nedaug. Yra vienaląsčių ir daugialąsčių dumblių.
Chlamidija, monada ir chlorelė. Maži vienaląsčiai dumbliai. Juos galima pamatyti tik mikroskopu. Jie turi apvalkalą, branduolį, citoplazmą, taurelės formos chromatoforą. Tačiau jie skiriasi esminėmis savybėmis.
Chlamidomonas
Aktyviai juda žvyneliais.
Yra raudonai šviesai jautri akis.
Yra pulsuojančių vakuolių.
Nelytinis dauginimasis judriomis sporomis su žvyneliais – zoosporomis.
Seksualinis procesas vyksta su dvipusių lytinių ląstelių pagalba.
Gyvena nedideliuose gėluose vandenyse.
Chlorella
Nejudėdamas.
Trūksta raudonos šviesai jautrios akies.
Pulsuojančių vakuolių nėra.
Nelytinis dauginimasis nejudrių sporų pagalba.
Seksualinis procesas nevyksta.
Daugiausia gyvena drėgnoje dirvoje ir medžių kamienuose.
Spirogyra. Labiausiai paplitę siūliniai žalieji dumbliai. Sudaro didžiąją dalį slidaus žaliojo purvo tvenkiniuose. Ląstelėje yra šios dalys: apvalkalas, padengtas gleivėmis; didelis branduolys su branduoliu (aiškiai matomas mikroskopu); citoplazma; chromatoforas vienos ar kelių juostelių, išdėstytų spirale, pavidalu; vakuolė, kuri užima didžiąją dalį ląstelės.
Vegetatyvinis dauginimasis spirogyra vyksta paprasčiausiai nutrūkus siūlams. Aseksualių sporų nėra. Seksualinis procesas yra konjugacija, t.y. normalių vegetatyvinių ląstelių, o ne specialių lytinių ląstelių susiliejimas. Konjugacijos žingsniai:
1 - du sriegiai yra išdėstyti lygiagrečiai;
2 - priešingos ląstelės sudaro ataugas viena link kitos, todėl susidaro kopėčių panašumas;
3 - apvalkalai procesų galuose ištirpsta;
4 - vienos iš ląstelių turinys patenka į priešingą ląstelę ir susilieja su jos turiniu, todėl susidaro zigota.
Cladophora yra išsišakojęs siūlas, kuris liesti nėra gleivingas, pritvirtintas prie substrato. Chromatoforas – tinklelio pavidalo. Kiekviena ląstelė turi daug branduolių (tai kartais randama dumbliuose, bet niekada aukštesniuose augaluose). Nelytinis ir lytinis dauginimasis vyksta maždaug kaip Chlamydomonas.
Taigi, keturių žaliųjų dumblių genčių požymiai yra tokie:
Jūros dumbliai. Kai kurie jūros dumbliai yra mikroskopiniai, dažnai vienaląsčiai. Daugelis jų gyvena paviršiniuose vandens sluoksniuose ir sudaro planktono dalį. Kiti gyvena dugne, daugiausia ant akmenų ir povandeninių uolienų, gana sekliame gylyje (150 - 200 m), t.y. daugiausia pajūrio zonoje.
Dumbliams reikia šviesos, todėl jie negali egzistuoti labai dideliame gylyje. Jų mažai ir kur vandenyje trūksta maistinių medžiagų. Didžiąją dugno dumblių dalį sudaro rudieji ir raudonieji dumbliai. Šių dumblių forma yra labai įvairi: krūmų, plokščių, virvelių pavidalu. Rudieji dumbliai yra rudos, rudos arba beveik juodos spalvos; raudona - rožinės spalvos, ryškiai - arba tamsiai raudona. Rudieji dumbliai pasiekia didžiausius dydžius tarp jūros dumblių. Tai visų pirma rudadumbliai arba jūros dumbliai.
Dumblių (talų) kūnas primena ilgą gana siaurą lapkočio lapą. Prie dugno prisitvirtina ataugos – šakniastiebiai. Kaip ir kiti dumbliai, rizoidai tarnauja tik tvirtinimui: vandenį sugeria visas paviršius. Laminaria siekia kelių metrų ilgį. Jo vidinė struktūra yra gana sudėtinga. Jis netgi turi sieto ląsteles, primenančias aukštesnių augalų sieto vamzdelius. Tačiau indų nėra, nes dumbliams jų nereikia. Kelpuose susidaro zoosporos, iš kurių išauga mikroskopinės ataugos su lytiniais organais. Taigi rudadumblių vystymosi ciklas kažkuo primena paparčius.
Fucus, taip pat rudasis dumblis, gyvena mūsų šiaurinių jūrų pakrantės zonoje. Fucus thallus yra stipriai išpjaustytas į diržą panašias skiltis. Jis yra daug mažesnis nei rudadumblių (iki 50 cm ilgio). Reprodukciniai organai formuojami specialiose talpyklose. Nelytinio dauginimosi sporų nėra. Jūros dumblių svarba iš esmės yra tokia:
planktoniniai dumbliai vaidina svarbų vaidmenį jūrų gyvūnų mityboje;
dugno dumblių tankiai suteikia prieglobstį žuvims ir kitiems gyvūnams;
dumblius ir kitus dumblius žmonės naudoja kaip maistą;
jodas ir agaras-agaras gaunami iš rudųjų ir raudonųjų dumblių;
Chlorela naudojama astronautikoje normaliai oro sudėčiai atkurti.
bryofitai
Bendrieji ženklai. Bryofitai – augalai, dažnai labai smulkūs, gana paprastos sandaros. Skirtingai nuo dumblių, jie dažniausiai turi lapus ir stiebus. Šaknų visada nėra; yra tik rizoidai. Lytiniai organai ir sporangijos yra daugialąsčiai. Vystymo ciklas labai ypatingas – iš zigotos tiesiai ant augalo išsivysto dėžutės su sporangijomis.
Briofitų struktūra. Žalios, arba skutimosi, samanos. Paskutinis epitetas yra sėkmingesnis, nes visi bryofitai yra žali augalai.
Tarp gegnių samanų vienas didžiausių atstovų yra gegutės linai. Jo stiebai siekia 20 cm ilgį (samanoms tai yra daug). Stiebas nešakotas, tankiai apaugęs siaurais lapeliais, šiek tiek primena tikrus linus (iš čia kilęs pavadinimas). Vietoj šaknų jie yra tiesiog išsidėstę rizoidai, besitęsiantys nuo stiebo apačios. Jie skirti ir pritvirtinti, ir sugerti vandenį (skirtingai nei dumbliai). Lyginant su dumbliais, brie samanos taip pat turi sudėtingą vidinę struktūrą. Pavyzdžiui, gegutės linai atrodo kaip epidermis ir laidus audinys.
Kukuškino linai yra dvinamis augalas: vyriški ir moteriški lytiniai organai yra ant skirtingų egzempliorių, šalia viršūnės. Vyriški reprodukciniai organai – anteridijos yra maišeliai, iš jų susidaro spermatozoidai. Moterų reprodukciniai organai – archegonijos yra panašios į kūgius su ilgais kakleliais. Jų siena susideda iš vieno ląstelių sluoksnio; Išsiplėtusioje kūgio dalyje yra kiaušialąstė. Tręšimui reikalingas lietus arba rasa. Tada spermatozoidai gali patekti ant archegoniumo ir per kaklą prasiskverbti į kiaušinėlį. Iš zigotos susidaro dėžutė ant ilgo kotelio. Dėžutė su dangteliu ir uždengta dangteliu. Viduje yra sankabos formos sporangiumas. Sporangijose susidaro sporos, kurios subrendusios iškrenta iš dėžutės. Norėdami tai padaryti, dangtelis turi nukristi, o sporangijos sienelė subyrėti. Aišku, kuo ilgesnis stiebas, tuo toliau gali plisti sporos. Sporos sudygsta, susidaro plonas žalias siūlas. Ant siūlo atsiranda pumpurai, iš kurių išauga samanų ūgliai.
Brie samanos yra labai paplitusios gamtoje. Jų galima rasti pelkėse, pievose, dykumose. Ypač daug jų pavėsinguose miškuose. Ne visi jie atrodo kaip gegutės linai. Daugelis stiebų yra stipriai šakoti, dažnai šliaužiantys. Yra daug samanų, kurių stiebai neviršija 2-3 cm.Dėžutės taip pat gali būti įvairių formų. Tačiau gyvenimo ciklas yra vienodas visiems.
Durpės, arba sfagnai, samanos. Durpynuose auga durpių samanos, kartu su spanguolėmis, mėlynėmis ir laukiniu rozmarinu. Tik labai nedaug augalų susitvarko su durpių samanomis. Jie visada pasirodo masėje, sudarydami vientisą kilimą. Sfagninių samanų stiebas šakojasi, suformuodamas trijų tipų šakas: vienos išsišakoja į šonus, kitos kabo žemyn, greta stiebo, o kitos sudaro savotišką galvą viršuje. Lapai yra labai maži (vos matomi plika akimi) ir susideda iš vieno ląstelių sluoksnio. Yra dviejų tipų ląstelės: dideli vandeningieji sluoksniai, skaidrūs, su spirale sustorėjusiomis sienelėmis ir siauri chlorofilą turintys, žali. Kiekvienas vandeningasis sluoksnis yra apsuptas kelių chlorofilą turinčių ląstelių. Vandeningojo sluoksnio ląstelės gali labai greitai absorbuotis puiki suma vandens (25 kartus daugiau nei sauso svorio) ir taip pat greitai jį numesite. Dėl šios savybės sfagnumas neturi ne tik šaknų, bet ir rizoidų (jam jų nereikia). Sfagninės samanos dauginasi taip pat, kaip ir brie samanos.
Sfagniniai augalai auga iš viršaus ir miršta iš apačios. Mirštančios apatinės dalys kartu su kitais augalais virsta durpėmis. Pastarasis susidaro nevisiškai irstant augalų dalims (nepakankamai deguonies). Durpės yra vertingas kuras. Tačiau daugeliu atvejų pelkių sausinimas yra nepageidautinas. Pirma, gali atsirasti klimato kaita; antra, sfagninėse pelkėse dažnai aptinkami reti augalai. Daugeliui sfagninių pelkių dabar suteiktas gamtos paminklų statusas.
paparčiai
Bendrieji ženklai. Paparčiai turi šaknis ir ūglius (stiebus su lapais). Jie dauginasi sporomis. Lyties organai formuojasi ant specialių smulkių augalų – ataugų.
Paparčių sandara. Paparčiai yra plačiai paplitę. Jie turi didelius, stipriai išpjaustytus lapus, besitęsiančius nuo šakniastiebių. Ant šakniastiebių taip pat susidaro atsitiktinės šaknys. Lapkočiai padengti rusvais žvynais. Jaunų lapų viršus sulankstytas į sraigę. Augimo procese sraigė išsivynioja, o lapas auga viršuje, kaip ūglis. Dėl šios savybės paparčio lapai kartais vadinami plokščiomis šakomis.
Paparčių dauginimasis. Apatinėje lapo pusėje (bet ne kiekviename) susidaro sporangijos, išsidėsčiusios grupelėmis ir dažnai padengtos šepetėliais arba lapo ašmenų kraštu. Plika akimi sunku pamatyti vieną sporangiją. Jo struktūra idealiai tinka sporoms išsklaidyti. Savo forma jis panašus į abipus išgaubtą lęšį. Sporangiumo sienelės sudarytos iš vieno sluoksnio ląstelių. Visi jie yra plonasieniai, išskyrus ląsteles, esančias palei keterą (žiedą). Šios ląstelės turi sustorėjusias vidines ir šonines sienas. Svarbu, kad žiedas užimtų ne visą keterą, o 2/3 jo, todėl lieka plonasienė kraigo dalis. Sporoms subrendus, sporos sienelė lūžta, o žiedas kaip spyruoklė išbarsto sporas. Iš sporos išauga mažytis augalas, prispaustas prie žemės širdies formos plokštelės pavidalu. Tai daigas. Turi rizoidų; apatinėje pusėje susidaro anteridijos ir archegonijos. Apvaisinimas vyksta kaip ir briofitai. Iš zigotos išsivysto embrionas, o vėliau – jaunas paparčio augalas.
Paparčių įvairovė. Paparčiai daugiausia yra miško augalai. Ypač daug jų drėgnuose atogrąžų miškuose. Daugumos jų lapai yra stipriai išpjaustyti, dažnai labai didelių dydžių. Tačiau yra daug paparčių su visais lapais. Vieni vijokliai laipiojančiais stiebais ar lapais, būna medžių pavidalo, 10 m ir aukštesniais kamienais. Tarp paparčių ypač daug epifitų, kurie nusėda ant medžių kamienų ir šakų. Vidutinio klimato platumose paparčių yra nedaug. Dažniausiai turime paparčio patiną, moterišką papartį (pavadinimai siekia senus laikus, kai dar nebuvo žinoma, kaip paparčiai dauginasi), braškę, strutį ir kai kuriuos kitus.
Asiūkliai ir klubinės samanos. Tai taip pat daugiametis žolinis sporinis augalas.
Jų savybės, palyginti su paparčiais, yra šios:
Buvęs paparčių žydėjimas. Paparčių grupėje priskaičiuojama 13 000 rūšių. Maždaug prieš 300 milijonų metų Žemėje nebuvo žydinčių augalų. Gimnosėklių jau atsirado, tačiau ypač svarbų vaidmenį atliko paparčiai. Daugelis iš jų buvo tikri medžiai, su kambiu, siekę 40 m aukštį, jų kamienai kartais būdavo ne mažesnio kaip 1 m skersmens. Vieni priminė iki milžiniškų dydžių padidintus asiūklius, kiti – klubines samanas. Vaistažoles taip pat reprezentavo tik paparčiai ir bryofitai. Klimatas buvo šiltas ir drėgnas, o apšvietimas ne toks intensyvus kaip dabar. Miškai dažnai buvo užpelkėję, nykstantys, medžiai įkrito į vandenį, buvo padengti dumblu. Palaipsniui kamienai buvo suspausti ir, neturint prieigos prie deguonies, virto anglimi – puikiu kuru.
Augalas yra visas organizmas
Augalų organai – tiek vegetatyviniai, tiek generaciniai – yra sudėtinguose santykiuose, užtikrinantys vieno organizmo gyvybę. Šaknys iš dirvožemio sugeria vandenį ir mineralines druskas, kurios būtinos normaliam visų gyvų ląstelių egzistavimui. Šaknyse susidaro organinės medžiagos: aminorūgštys, vitaminai, hormonai, fermentai ir kiti junginiai, be kurių neįmanoma organizmo gyvybė. Kai kurie iš jų patenka į chlorofilo susidarymą lapuose. Be chlorofilo fotosintezė nevyksta. Fotosintezei reikalingas vanduo, kuris taip pat patenka į žaliąsias lapo ląsteles iš šaknų.
Didelį kiekį vandens išgarina antžeminiai organai ir taip augalas apsisaugo nuo perkaitimo. Šaknys aprūpina ūglius vandeniu.
Savo ruožtu šaknų ląstelėse galima įvairių gyvybiškai svarbių junginių sintezė, kai jie iš lapų gauna organinių medžiagų. Tik ląstelėse su chloroplastais organinės medžiagos susidaro iš neorganinių medžiagų – vandens ir anglies dioksido. Fotosintezės produktai būtini šaknims augti ir šakotis.
Taigi, tik esant glaudžiui antžeminių ir požeminių vegetatyvinių organų ryšiui, įmanoma organizmo gyvybė.
Žydėjimas, vaisių ir sėklų nokinimas taip pat neįmanomas, jei gimdos organai nėra aprūpinti visomis jiems reikalingomis medžiagomis. Šias medžiagas jiems tiekia vegetatyviniai organai.
Savo ruožtu generatyviniai organai veikia vegetatyvinių organų gyvybinę veiklą. Taigi šaknų darbas priklauso ne tik nuo oro mitybos organų, lapų, bet ir nuo generatyvinių organų. Eksperimentais buvo įrodyta, kad pašalinus kiaušides iš kviečių žiedų eilės arba užtemdžius ausis, pastebimai sumažėjo azoto tiekimas iš šaknų į antžeminę augalo dalį.
Šie pavyzdžiai rodo, kad augalo organizmas yra viena ir vientisa sistema. Šioje sistemoje funkcijos skirstomos tarp atskirų įstaigų, tačiau jų veikla yra glaudžiai tarpusavyje susijusi.
Dienos metu ar ištisus metus temperatūra keičiasi judant nuo pusiaujo į ašigalius ir kt. Gyvūnams egzistavimas gali būti laikomas normaliu, jei pasikeičia aplinkos veiksnys neviršija didžiausių ir mažiausių verčių, ty tolerancijos ribų.
Tarp verčių, ribojančių gyvūno egzistavimo galimybę, yra jam palankiausia, bet gana siaura optimali zona. Tarp optimalios zonos ir maksimumo arba minimumo yra pesimumo zona, kurioje nepalankios sąlygos veikia organizmą.
Optimumo ir pesimumo zonos nustatomos pagal individų skaičių ploto vienete, tai yra pagal rūšių populiacijų tankį.
1 apibrėžimas
Rūšies ekologinis valentingumas – tai tam tikros rūšies gebėjimas apsigyventi skirtinga aplinka, kuriai būdingi įvairūs aplinkos veiksnių pokyčiai.
Stenobiont vaizdas yra mažo ekologinio valentingumo rūšis, prisitaikiusi ištverti tik ribotus aplinkos veiksnių svyravimus. Sausumos atstovai: humanoidinė šimpanzė, gyvenanti atogrąžų miškuose, vidutinio klimato juostos spygliuočių miško gyventoja - kurtinys. Vandens aplinkos stenobiontai: rifus formuojantys koralai, kurių normaliam augimui ir vystymuisi reikalinga griežtai apibrėžta jūros vandens temperatūra, apšvietimo laipsnis ir druskingumas. Šie koralai gyvena tropinėje vandenynų zonoje.
eurybiontų rūšis- tai rūšis, galinti apsigyventi buveinėse su besikeičiančiomis sąlygomis arba įvairiose buveinėse (lapė randama kalnuose ir lygumose, stepėse ir miškuose, keliose kraštovaizdžio ir klimato zonose; banginiai gali egzistuoti įvairiose platumose ).
Nurodant santykinį tolerancijos laipsnį, atitinkami terminai vartojami su priešdėliu "steno" - siauras arba "evry" - platus:
- santykinės temperatūros – stenoterminė arba euriterminė;
- dieta – stenofagai arba eurifagai;
- vandens druskingumas – stenohalinas arba eurihalinas;
- buveinės atžvilgiu – stenoek ir euryek ir kt.
Gyvūnų pasiskirstymo ir egzistavimo jūros aplinkoje sąlygos
Jūrose ir vandenynuose yra turtingiausia fauna, kuri sudaro 64% visų gyvūnų rūšių. Daugelio rūšių gyvūnai gali gyventi tik jūroje: brachiopodai, koralų polipai, galvakojai ir šoniniai nerviniai moliuskai, gaubtagyviai, akraniatai, kempinės, nemerteanai, daugiasluoksniai žiedai ir kt.
Jūrų organizmus ypač veikia:
- slėgis;
- tankis;
- saulės spindulių įsiskverbimo gylis;
- druskos tirpimas ir dujų kiekis;
- šilumos paskirstymas;
- srovės.
Jūros aplinkoje yra dvi biotopų grupės:
- Pelaginis – jūros vandens storis, kuriame gyvybė nesusijusi su kietu substratu (krantu, dugnu). Pelaginių žuvų atstovai plaukia aktyviai arba pasyviai (nektonas, planktonas, banginių šeimos gyvūnai, žuvys, moliuskai (kalmarai), stambieji galvakojai).
- Bentalis – visas dugno ir pakrantės paviršius. Bentalams būdingas vertikalus skrodimas, jie išskiria pakrantės ir bedugnės zonas. Tarp šių zonų nėra ryškios ribos, jas skiria batialinė zona.
Pajūrio zona apima kontinentinio šelfo paviršių nuo purslų zonos iki žemyninio šlaito pradžios. Zona pasižymi labai įvairiais biotopais ir maisto gausa.
bedugnės zona susideda iš jūros dugno ir šalia esančio vandens sluoksnio nuo žemyninio šlaito krašto. Monotoniškiausia buveinė planetoje. Maisto ištekliai yra riboti, juos daugiausia sudaro detritas. Pagrindiniai dideliame gylyje gyvenantys gyventojai yra dygiaodžiai. Dugne gyvena filtrų lesyklėlės: kirminai, jūrų anemonai, moliuskai, jūros vorai pantopodai, krabai ir kiti gyvūnai. Yra daug relikvijų - chimeros žuvys, jūros lelijos ir kai kurie kiti atstovai.
Gyvūnų pasiskirstymo ir egzistavimo gėluose vandenyse sąlygos
Gėlo vandens telkiniai yra atskirti ir tarp juose gyvenančių gyvūnų nėra tiesioginių ryšių. Gėlo vandens telkiniuose galvakojų, dygiaodžių, daugiasluoksnių žiedų aptikti nepavyksta, tačiau gali pasitaikyti koelenteratų ir kempinių atstovų. Amfibijos gyvūnai siejami tik su gėlo vandens telkiniais.
Gėlavandenė fauna yra iš jūros, todėl organizmų pasiskirstymą įtakoja bendrieji aplinkos veiksniai:
- Rezervuarų druskingumas. Kai kurios gyvūnų rūšys – eurihaliniai organizmai – prisitaikė prie vandens druskingumo svyravimų. Stenohaliniai organizmai yra griežtai selektyvūs druskingumo laipsniui.
- Vandens kietumas. Taigi, briozai, kempinės, kai kurios vėžių rūšys gali gyventi tik minkštame vandenyje.
- Humuso ir ištirpusio deguonies kiekis. Rezervuaruose, kuriuose gausu humuso, gyvų organizmų yra nedaug. Dėl organinių medžiagų irimo žūsta žuvys, o tai ypač pavojinga stenoksibiontams.
- Temperatūra. Dauguma vidaus vandenų atstovų priskiriami euriterminiams organizmams.
- srovės. Gėlo vandens telkinių atstovai jautrūs vandens judėjimo greičiui, jie skirstomi į reofilus – tekančių vandenų gyventojus ir limnofilus – stovinčio vandens gyventojus.
Sausumos gyvūnų paplitimo ir egzistavimo sąlygos
Sausumoje yra dideli visų aplinkos veiksnių svyravimai.
Pagrindiniai veiksniai, lemiantys sausumos gyvūnų pasiskirstymą:
- Oro ir dirvožemio drėgmė. Yra higrofilai – drėgmę mėgstantys gyvūnai ir kserofilai – sausamėgiai, yra organizmų, kurių egzistavimas nepriklauso nuo drėgmės.
- Oro judėjimas. Prieinamumas stiprus vėjas(salos, jūros pakrantės) atbaido skraidančius vabzdžius, o už jų išnyksta vabzdžiaėdžiai gyvūnai.
- Temperatūra. Kiekviena rūšis turi savo temperatūros diapazoną – rūšies temperatūros optimalumą, kuris užtikrina palankų egzistavimą. Euriteminės rūšys daugiausia gyvena vidutinio klimato juostose, stenoterminės rūšys skirstomos į termofilines (politermines) – mylinčias šilumą ir termofobines (oligotermines) – mėgstančias šaltį.
- Saulės radiacija.Šviesa daro įtaką dienos ir nakties gyvūnų formų egzistavimui. Didelę įtaką turi šviesaus paros valandų trukmė. Fotoperiodizmo reiškinys nulemia griežtai apibrėžto vabzdžių kartų skaičiaus vystymąsi per metus, teritorijų išplėtimą į kitas platumos zonas ir kt.
- Substrato pobūdis. Gyvūnų pasiskirstymas priklauso nuo dirvožemio druskingumo. Nariuotakojai yra jautriausi druskingumui. Pavyzdžiui, Bledius genties vabalai aptinkami tik druskingose dirvose. Dirvožemio chemija gali turėti netiesioginį poveikį gyvūnams per pašarinius augalus.
- Augalinė danga lemia biocenozės pobūdį ir veikia kaip jos indikatorius. Pavyzdžiui, Šiaurės spygliuočių miškuose, kur auga laukiniai rozmarinai, bruknės, žaliosios samanos, galima sutikti maskvietines zyles, kurtinius, mišrūnus, veržles, sabalus, burundukus, lūšis.
Arba dirbtiniai: tvenkinys, rezervuaras, kanalas (4-6 pav.).

Ryžiai. 5. Rezervuaras ()

Kad ir koks būtų rezervuaras, natūralus, dirbtinis, jis puošia mūsų kraštą, džiugina mus savo grožiu. Į šviežius rezervuarus mes imame vandenį, be kurio negalime išsiversti nei namuose, nei darbe. Plaukiame rezervuaruose, deginamės šalia jų, keliaujame vandeniu laivais, vežame prekes. Vandens telkinių reikšmė gamtoje didelė. Gėlas vanduo yra esminė sąlygažmonių egzistavimo Žemėje ir gyvūnams, gyvenantiems vandenyje, namai taip pat yra vieninteliai namai. Vanduo turi viską, ko reikia gyvybei: šviesą, šilumą, orą ir ištirpusius mineralus.
Kokie augalai auga ir kokie gyvūnai gyvena gėlame vandenyje? Patekę į rezervuarą šiltuoju metų laiku, galėjote stebėti tik tuos jo gyventojus, kurie gyvena paviršiuje. Tačiau gyvybė rezervuare yra visur: ir pakrantėje, ir paviršiuje, ir vandens storymėje, pačiame dugne ir apačioje. Ant rezervuarų krantų galima pamatyti nendrių, nendrių, kačių, strėlių antgalių lapus ir stiebus. Mažas gylis leidžia šiems augalams prisitvirtinti prie rezervuaro dugno. Daug didesniame gylyje auga baltoji vandens lelija ir geltona ankštis (7, 8 pav.). Ant lygus paviršius vandenys plūduriuoja savo žiedais ir plačiais lapais.

Ryžiai. 7. Baltoji vandens lelija ()

Ryžiai. 8. Geltona kapsulė ()
Kaip šiems augalams pavyko prisitaikyti prie gyvenimo labai drėgnoje dirvoje, kur beveik nėra deguonies? Atsižvelgdami į nendrių, nendrių, kačių stiebų atkarpą, pamatysime oro kanalus, kurie eina šių augalų stiebuose (9, 10 pav.).

Ryžiai. 9. Nendrė ()

Vandens augalų lapuose ir šaknyse yra oro kanalų. Baltojoje vandens lelijoje ir geltonoje kapsulėje lapų stiebai ir žiedkočiai, ant kurių sėdi gėlės, taip pat yra perverti oro kanalais, pro kuriuos prasiskverbia kvėpavimui reikalingas deguonis. Nuskindamas gėlę žmogus kenkia visam augalui. Vanduo pradeda prasiskverbti į augalą plyšimo vietoje, o tai veda prie povandeninės dalies irimo ir galiausiai viso augalo žūties.
Ančiuko žolė mažų žalių plokštelių pavidalu taip pat plūduriuoja rezervuaro paviršiuje, bet prie dugno neprisiriša savo šaknimis, o vandens storymėje yra patys mažiausi žalieji dumbliai, juos galima pamatyti tik pro mikroskopą. Tačiau jų buvimas išduoda vandens spalvą. Kai rezervuare jų yra daug, vandens spalva tampa žalia.
Kokį vaidmenį augalai vaidina daugelio vandens telkinių gyventojų gyvenime? Pirma, žalieji augalai, veikiami saulės spindulių, paima iš oro anglies dvideginį, o į vandenį išskiria deguonį, reikalingą visų gyvūnų kvėpavimui. Antra, rezervuaro tankmėje prieglobstį ir maistą randa paukščiai, varliagyviai, vabzdžiai ir jų lervos, žuvys. Gyvūnų rezervuaruose yra visur: paviršiuje ir vandens storymėje, krante, dugne, ant vandens augalai. Pagrindinės gyvūnų ir augalų sąsajos yra maistas. Štai vandens strideriai (11 pav.) greitai laksto vandens paviršiumi ir medžioja uodus bei kitus smulkius gyvūnus.

Ryžiai. 11. Vandens čiuožykla ()
Jų ilgos kojos iš apačios padengtos riebalais, todėl vanduo jas laiko. O sraigės gyvena ant vandens augalų: kūdros sraigės ir spiralės (12, 13 pav.).

Ryžiai. 12. Prudovikas ()

Be ko upė negali gyventi? Labai maži rezervuarų vėžiagyviai, dafnijos ir ciklopai, gyvena ir žiemoja vandenyje. Jų reikšmė kiek didesnė už knygoje esantį kablelį (14, 15 pav.).
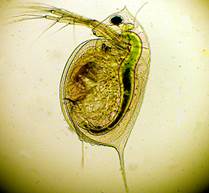
Įspūdingiausias dafnijos dalykas yra ilgi ūsai. Jie mojuoja ūsais, smarkiai nukrenta, atsistumia nuo vandens ir šokinėja. Ciklopas turi nesuporuotą priekinį oką, nuo kurio ir gavo savo pavadinimą.

Upė negali gyventi be vėžiagyvių, nes jie išvalo vandenį nuo bakterijų, žaliųjų dumblių ir mažiausių akiai nematomų gyvūnų, jei ne vėžiagyviai, upė jais greitai išsilietų. Dafnijos ir ciklopai, kaip ir kiti upės gyventojai, minta šiais organizmais ir taip valo vandenį. Jie patys tarnauja kaip maistas žuvų mailiui, moliuskams, buožgalviams, vabzdžių lervoms.
Ar kas nors gyvena upėje be galvos? Tai moliuskai, bedantys ir miežiai (16 pav.).

Ryžiai. 16 moliuskų ()
Iš pradžių apvalkalas, susidedantis iš dviejų išilginių plokštelių, gulės nejudėdamas, tada jo vožtuvai šiek tiek atsidarys ir iš jo išlįs koja, nei bedantis, nei miežiai neturi galvos. Bedantis ištiesia koją ir įkiša ją į smėlį, kiautas pasislinks. Bedantė pajudės 2-3 centimetrus, pailsės – ir vėl kelyje. Taigi jis keliauja upės dugnu. Be dantų maistą ir orą gauna tiesiai iš vandens. Jis atidaro kriauklės apvalkalus ir pradeda traukti vandenį, tada jį išmeta. Vanduo pilna smulkiausių gyvūnų, jie krenta į kriauklę, todėl bedantukai sulaiko juos specialiais prietaisais. Bedantis kvėpuoja ir valgo, o tuo pačiu valo vandenį. Ir perlų juosta taip pat veikia. Kiekvienas išvalo maždaug 40-50 litrų vandens per dieną. Moliuskus, vabzdžių lervas, buožgalvius minta žuvys, gandrai, smėlinukai, antys. Plaukiantis vabalas grobia kitus vabzdžius, taip pat kirmėles, sraiges ir buožgalvius. Varlės maitinasi vandens telkinių pakrančių dalyse, daugiausia skraidančiais vabzdžiais, o pačios yra maistas tritonams ir plėšriosioms žuvims, ešeriams ir lydekoms. Garniai, kirai, karaliukai medžioja žuvis ir tritonus.
Pagrindinis vėžio maistas yra daržovės. Tačiau jis noriai valgo gyvūnus, taip pat ir negyvų gyvūnų palaikus. Todėl vėžiai dažnai vadinami rezervuarų tvarkdariais (17 pav.).

Vėžiai per savo gyvenimą keičia savo apvalkalą. Jutimo organai vėžiai puikiai išsivysčiusios, akys nustumtos į priekį ant plonų kotelių ir susideda iš daugybės, 3000, mažyčių akių. Trumpa antenų pora yra uoslės organai, o ilgosios – lytėjimo organai. Jei plėšrūnas sugriebia vėžį už nagų, vėžys jį nulaužia ir pasislepia duobėje. Pamestas letenas ataugs. Vėžiai labai jautrūs vandens taršai, todėl jų aptikimo vietose kalbama apie ekologinę vandens telkinių švarą.
Prie upės galima pamatyti įvairių laumžirgių: gražuolį, strėlę, lutką, jie visą laiką gyvena prie upės (18 pav.).

Ryžiai. 18. Laumžirgis ()
Visiems laumžirgiams reikia vandens, nes tik ten gali gyventi jų lervos. Lervos nepanašios į suaugusius laumžirgius, tik jų akys vienodos. Kiekviena akis susideda iš beveik 30 000 mažyčių okelių (19 pav.).

Ryžiai. 19. Laumžirgio lerva ()
Abi akys yra išgaubtos, dėl to laumžirgis gali vienu metu žiūrėti į visas puses (20 pav.). Visi laumžirgiai yra plėšrūnai, jie medžioja ore, skraidydami gaudo vabzdžius.

Ryžiai. 20. Laumžirgio akys ()
Laumžirgio lerva, stebėjusi grobį, meta į priekį stipriai pailgą apatinę lūpą. Dažniausiai lūpos yra sulenktos ir dengia galvą kaip kaukė. Lerva įsiurbia vandenį į didelį raumeninį maišelį kūno viduje, o paskui jėga jį išmeta. Pasirodo, vandens šūvis. Po metų, o kai kurios ir po 3, lervos patenka į paviršių, lervos oda plyšta, nuo jos atsiranda laumžirgis. Kelias valandas sėdės, išskleis sparnus ir išskris.
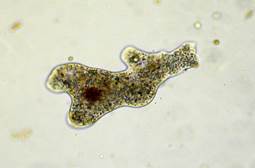
Kas gyvena vandens laše? Jei pažiūrėsite pro mikroskopą, pamatysite nuostabus pasaulis neįprasti padarai. Štai beveik skaidrus gumulas, kuris visą laiką keičiasi – tai ameba (21 pav.).
Kiti padarai primena mažytes šlepetes, kaip jos vadinamos. Batų korpusas padengtas blakstienomis, kiekvienas meistriškai valdo šias blakstienas ir greitai plaukia (22 pav.).
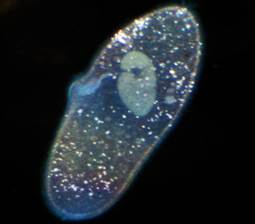
Ryžiai. 22. Batas ()
Trimitininkai – gražiausi lašo gyventojai, mėlyni, žali, panašūs į uolienų žiedus (23 pav.).
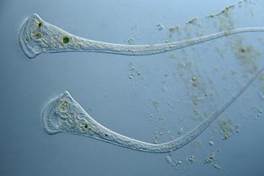
Trimitininkai juda lėtai ir tik į priekį. Jei kas nors juos gąsdina, jie susitraukia ir primena kamuoliukus. Amebos, šlepetės ir trimitininkai yra vienaląsčiai organizmai, mintantys bakterijomis.
Plėšrūnai taip pat gyvena vandens laše. Tai didinis (24 pav.).
Nors jis mažesnis už batą, ne tik drąsiai puola ją, bet ir praryja visą, ištinusią kaip balionas.
AT gėlo vandens augalai, gyvūnai, bakterijos gyvena kartu, visi jie puikiai prisitaikę gyventi vandenyje ir yra tarpusavyje susiję mitybos grandinėmis. Žudami augalai ir gyvūnai kaupiasi vandens telkinių dugne, juos sunaikina bakterijos ir virsta vandenyje tirpstančiomis druskomis, kurias naudoja kiti gyvūnai. Vandens telkinys yra natūrali bendrija.
Šiandien pamokoje įgijote naują idėją apie gėlo vandens telkinį kaip gėlo vandens bendruomenę ir susipažinote su jo gyventojais.
Bibliografija
- Vachruševas A.A., Danilovas D.D. Pasaulis 3. - M.: Ballas.
- Dmitrieva N.Ya., Kazakovas A.N. Pasaulis aplink 3. - M .: leidykla "Fedorov".
- Plešakovas A.A. Pasaulis aplink 3. - M .: Švietimas.
- Makuha.ru ().
- Youtube.com ().
- sbio.info().
Namų darbai
- Kokius gėlo vandens telkinius žinote?
- Kokius gyvūnus galima rasti vandens telkiniuose?
- Kodėl sakoma, kad vandens telkinys yra natūrali bendrija?
Rudieji ir raudonieji dumbliai, bendrosios charakteristikos, morfologija, fiziologijos pagrindai, gyvavimo ciklų specifika, taksonomija, vaidmuo biosferoje ir žmogaus gyvenime.
Raudonieji dumbliai arba Bagryanka. Didžioji dauguma raudonųjų dumblių yra jūrų augalija ir gyvūnija bet jų randama ir gėluose vandenyse.
Bagryanki - daugialąsčiai organizmai, turinčios siūlinę ir sluoksninę struktūrą, ir tik kelios, pačios primityviausios, yra vienaląstės arba kolonijinės formos. Daugelis iš jų yra dideli augalai, kurių ilgis siekia keliasdešimt centimetrų, tačiau tarp jų yra daug mikroskopinių formų. Pagal formą raudondumbliai yra gijų, krūmų, plokščių, plutų, koralų ir kt. Tvirtinimo organai yra šakniastiebiai, čiulptukai ir padai. Jų talio spalva svyruoja nuo ryškiai raudonos iki melsvai žalios ir geltonos. Taip yra dėl to, kad lameliniuose chromatoforuose, be chlorofilo, yra raudonojo pigmento fikoeritrino ir mėlynojo fikocianino.
Jie dauginasi nelytiškai (sporomis) ir lytiškai.
Rudieji dumbliai. Dažnas rudųjų dumblių, gyvenančių visose Žemės rutulio jūrose, išorinis požymis yra gelsvai ruda jų talio spalva, kurią lemia jų ląstelėse esantys geltoni ir rudi pigmentai (karotinai ir ksantofilai).
Rudųjų dumblių talas primena išsišakojusius krūmus, lėkštes, juosteles, kompleksiškai suskaidytus į stiebo ir lapo formos organus. Talio dydis svyruoja nuo kelių centimetrų iki 60-100 m. Talis auga dėl tarpkalinio (tarpkalinio) augimo arba dėl nuolatinio viršūninės ląstelės dalijimosi. Pritvirtinimui prie žemės naudojami rizoidai arba bazinis diskas – disko formos atauga talio apačioje.
Pagal morfologinę ir anatominę talijos diferenciaciją rudųjų dumblių yra daugiau aukštas lygis nei visos kitos grupės. Tarp jų nežinomos nei vienaląstės, nei kolonijinės formos, nei paprasto nešakoto gijos pavidalo talis. Daugumoje atstovų talis turi klaidingą arba tikrą audinių struktūrą (išskiriami asimiliaciniai, kaupiamieji, laidūs audiniai).
Rudieji dumbliai dauginasi nelytiškai (talais arba sporomis) ir lytiškai. Lytinio dauginimosi metu zigota sudygsta į naują augalą be ramybės periodo.
Daugumos rudųjų ir raudonųjų dumblių vystymosi cikle vyksta reguliarus kartų kaitaliojimas – gametofitas ir sporofitas.
Stambių rudųjų dumblių tirščiai kartais nusidriekia dešimtis kilometrų, formuodami originalius povandeninius miškus ir pievas 40-100 m ar daugiau gylyje. Jie yra daugelio gyvūnų rūšių maistas, veisimosi ir prieglaudos, mikroorganizmų ir makroorganizmų substratas, vienas pagrindinių vandens telkinių organinių medžiagų šaltinių.
Dumblių vertė. Visur esantis dumblių paplitimas lemia didelę jų reikšmę biosferoje ir žmogaus ūkinėje veikloje. Dėl savo gebėjimo fotosintezuoti jie yra pagrindiniai didžiulio kiekio organinių medžiagų gamintojai vandens telkiniuose, kuriuos plačiai naudoja gyvūnai ir žmonės.
Sugerdami iš vandens anglies dioksidą, dumbliai prisotina jį deguonimi, reikalingu visiems gyviems organizmams. Jų vaidmuo biologiniame medžiagų cikle yra didelis.
Istorinėje ir geologinėje praeityje dumbliai dalyvavo formuojant uolienas ir kreidos uolienas, kalkakmenis, rifus, specialias anglies rūšis ir daugybę naftingųjų skalūnų. Jie buvo žemėje gyvenusių augalų protėviai.
Dumbliai plačiai naudojami šalies ekonomikoje, įskaitant maisto, farmacijos ir kvepalų pramonę. Jie auginami dideliu mastu augaluose po atviras dangus siekiant gauti biomasę kaip papildomą baltymų, vitaminų ir biostimuliatorių šaltinį gyvulininkystei. Taigi nustatyta, kad jūros dumbliuose yra vitaminų A, B1, B2, B12, C ir D, jodo junginių, bromo ir kt.
Daugelis dumblių naudojami žmonių maistui. Didžiausio pripažinimo sulaukė jūros dumbliai (kai kurios rudadumblių ir porfyro rūšys), kurie, be to, naudojami kaip gydomoji ir profilaktinė priemonė nuo virškinamojo trakto sutrikimų, sklerozės, strumos, rachito ir kitų ligų.
Dumbliai tarnauja kaip žaliava daugybei vertingų medžiagų gauti: alkoholiai, lakas, amoniakas, organinės rūgštys, jodas, bromas (rudas); agaras-agaras (raudonas). Agar-agaras buvo plačiai pritaikytas biotechnologijoje kaip kieta terpė bakterijoms, dumbliams, grybams auginti. Dideliais kiekiais iš jo gaminamas marmeladas, zefyras, ledai ir kt.
Žemės ūkyje dumbliai naudojami kaip organinės trąšos kai kuriems augalams ir kaip pašarų priedas naminių gyvūnų racione.
Kai kurie dumbliai (pavyzdžiui, chlorelė, scenedesmus ir kt.) gali kaupti radionuklidus, kurie gali būti naudojami papildomai valyti žemo aktyvumo nuotekas iš atominių elektrinių.
Žalieji dumbliai. Bendrosios charakteristikos, morfologija, fiziologijos pagrindai, gyvavimo ciklų specifika. Struktūrų raida ir dauginimosi būdai. Sistematika, pagrindiniai atstovai. Žalieji dumbliai kaip aukštesniųjų augalų protėviai. Vaidmuo biosferoje ir žmogaus gyvenime.
Žalieji dumbliai yra plačiausiai iš visų dumblių padalinių, kuriuose, įvairiais skaičiavimais, yra nuo 4 iki 13-20 tūkstančių rūšių. Visi jie turi žalia spalva thalli, kuris atsiranda dėl to, kad chloroplastuose vyrauja chlorofilas a ir b, palyginti su kitais pigmentais. Kai kurių žaliųjų dumblių (Chlamydomonas, Trentepolia, Hematococcus) atstovų ląstelės yra raudonos spalvos arba oranžinė spalva a, kuri yra susijusi su karotinoidinių pigmentų ir jų darinių kaupimu už chloroplasto ribų.
Morfologiškai jie yra labai įvairūs. Tarp žaliųjų dumblių yra vienaląsčių, kolonijinių, daugialąsčių ir neląsčių atstovų, aktyviai judančių ir nejudančių, prisirišusių ir laisvai gyvenančių. Jų dydžių diapazonas taip pat itin didelis – nuo kelių mikrometrų (kurių dydžiu prilygsta bakterijų ląstelėms) iki 1–2 metrų.
Ląstelės yra vienabranduolinės arba daugiabranduolinės, turinčios vieną ar daugiau chromatoforų, kuriuose yra chlorofilo ir karotinoidų. Chloroplastai yra padengti dviem membranomis ir paprastai turi stigmą arba akutę, filtrą, kuris praleidžia mėlyną ir žalią šviesą į fotoreceptorių. Akis susideda iš kelių lipidų rutuliukų eilių. Tilakoidai – dariniai, kuriuose yra lokalizuoti fotosintetiniai pigmentai – surenkami į krūvas (lameles) po 2–6. Žvaigždžių pereinamojoje zonoje yra žvaigždžių darinys. Paprastai yra dvi žvyneliai. Pagrindinė ląstelės sienelės sudedamoji dalis yra celiuliozė.
Chlorofitai maitinasi įvairiais būdais: fototrofiškai, miksotrofiškai ir heterotrofiškai. Žaliųjų dumblių rezervinis polisacharidas – krakmolas – nusėda chloroplasto viduje. Chlorofitai taip pat gali kaupti lipidus, kurie lašelių pavidalu nusėda chloroplasto stromoje ir citoplazmoje.
Daugialąsčiai taliai yra siūliški, vamzdiški, sluoksniniai, krūminiai arba skirtingos struktūros ir įvairių formų. Iš žinomų žaliųjų dumblių talijos organizavimo tipų nėra tik ameboido.
Jie plačiai paplitę gėluose ir jūros vandenyse, dirvožemyje ir sausumos buveinėse (ant dirvožemio, uolų, medžių žievės, namų sienų ir kt.). Jūrose paplitusi apie 1/10 visų rūšių, kurios dažniausiai auga viršutiniuose vandens sluoksniuose iki 20 m. Tarp jų yra planktoninės, perifitoninės ir bentosinės formos. Kitaip tariant, žalieji dumbliai įvaldė tris pagrindines gyvų organizmų buveines: vandenį – žemę – orą.
Žalieji dumbliai turi teigiamą (judėjimas link šviesos šaltinio) ir neigiamą (judėjimas nuo ryškios šviesos šaltinio) fototaksį. Be šviesos intensyvumo, temperatūra taip pat turi įtakos fototaksiui. Teigiamas fototaksas esant 160°C temperatūrai rodo Hematococcus, Ulotrix, Ulva ir genčių zoosporas. tam tikrų tipų desmidiniai dumbliai, kuriuose ląstelių judėjimas vyksta dėl gleivių išskyrimo per apvalkalo poras.
Reprodukcija.Žaliesiems dumbliams būdingi visi žinomi dauginimosi būdai: vegetatyvinis, nelytinis ir seksualinis.
Vegetatyvinis vienaląsčių formų dauginimasis vyksta ląstelėms dalijantis per pusę. Kolonijinės ir daugialąstės chlorofito formos dauginasi kūno dalimis (talas arba talis).
Nelytinis žaliųjų dumblių dauginimasis yra plačiai paplitęs. Jį dažniau atlieka judrios zoosporos, rečiau – nejudrios aplanosporos ir hipnosporos. Ląstelės, kuriose susidaro sporos (sporangijos), dažniausiai niekuo nesiskiria nuo likusių talio vegetatyvinių ląstelių, rečiau būna kitokios formos ir didesnių dydžių. Susidarančios zoosporos gali būti plikos arba padengtos standžia ląstelės sienele. Žvynelių skaičius zoosporose svyruoja nuo 2 iki 120. Zoosporos būna įvairių formų: sferinės, elipsoidinės arba kriaušės formos, vienabranduolės, neturinčios atskiros membranos, priekyje turi 2–4 žiuželius, smailesniu galu ir chloroplastu ties išplėstas užpakalinis galas. Paprastai jie turi pulsuojančias vakuoles ir stigmą. Zoosporos susidaro pavieniui arba, dažniau, tarp kelių vidinių motininės ląstelės turinio, išeina į lauką per apvalkale susidariusią apvalią ar plyšinę skylutę, rečiau dėl jo bendrų gleivių. Išėjimo iš motininės ląstelės momentu zoosporas kartais gaubia plona gleivėta šlapimo pūslė, kuri netrukus išsiskleidžia (Ulotrix gentis).
Daugelyje rūšių vietoj zoosporų arba kartu su jomis susidaro nejudančios sporos – aplanosporos. Aplanosporos yra nelytinės sporos, neturinčios žvynelių, bet turinčios susitraukiančias vakuoles. Aplanosporos laikomos ląstelėmis, kuriose sustabdomas tolesnis zoosporų vystymasis. Jie taip pat atsiranda iš ląstelės protoplasto, tarp vienos ar kelių, bet negamina žvynelių, o įgavę sferinę formą, apsirengia savo membrana, kurios formavime motininės ląstelės membrana nedalyvauja. . Aplanosporos išsiskiria dėl motininių ląstelių membranų plyšimo ar gleivių ir sudygsta po ramybės periodo. Aplanosporos su labai storais apvalkalais vadinamos hipnosporomis. Paprastai jie perima ramybės stadijos funkciją. Autosporose, kurios yra mažesnės nejudrių vegetatyvinių ląstelių kopijos, trūksta susitraukiančių vakuolių. Autosporų susidarymas koreliuoja su sausumos sąlygų užkariavimu, kai vandens ne visada gali būti pakankamai.
Lytinį dauginimąsi vykdo lytinės ląstelės, kurios atsiranda nepakitusiose, šiek tiek pakitusiose arba reikšmingai transformuotose ląstelėse – gametangijoje. Judrios monadinės struktūros lytinės ląstelės, biflageliuotos. Seksualinis procesas žaliuosiuose dumbliuose vaizduojamas įvairiomis formomis: hologamija, konjugacija, izogamija, heterogamija, oogamija. Esant izogamijai, gametos yra morfologiškai visiškai panašios viena į kitą ir skirtumai tarp jų yra grynai fiziologiniai. Zigota yra padengta storu kiautu, dažnai su skulptūrinėmis išaugomis, didelis skaičius atsargines medžiagas ir sudygsta iš karto arba praėjus tam tikram ramybės periodui. Dygimo metu daugumos rūšių zigotos turinys yra padalintas į keturias dalis, kurios išlenda iš apvalkalo ir išdygsta į naujus individus. Daug rečiau gametos į naują organizmą išsivysto be susiliejimo, savaime, nesusiformavus zigotai. Toks dauginimasis vadinamas partenogeneze, o iš atskirų lytinių ląstelių susidarančios sporos – partenosporomis.
Esant heterogamijai, abi gametos skiriasi dydžiu, o kartais ir forma. Didesnės lytinės ląstelės, dažnai mažiau judrios, laikomos moteriškomis, mažesnės ir judresnės – vyriškomis. Kai kuriais atvejais šie skirtumai yra nedideli, o tada jie tiesiog kalba apie heterogamiją, o kitais - labai reikšmingi.
Jei patelės lytinė ląstelė nejuda ir panašesnė į kiaušinėlį, tai mobilusis patinas tampa spermatozoidu, o lytinis procesas vadinamas oogamija. Gametangijos, kuriose gaminasi kiaušinėliai, vadinamos oogonia; jie skiriasi nuo vegetatyvinių ląstelių forma ir dydžiu. Gametangija, kurioje gaminasi spermatozoidai, vadinama anteridijomis. Zigota, susidaranti spermatozoidui apvaisinant kiaušinėlį, sudaro storą apvalkalą ir vadinama oospora.
Tipiškos oogamijos atveju kiaušinėliai yra dideli, nejudrūs ir dažniausiai vystosi po vieną oogonijoje, spermatozoidai yra maži, judrūs, daug jų susidaro anteridiume. Oogonia ir antheridia gali išsivystyti ant to paties individo, tokiu atveju dumbliai yra vienanamiai; jei jie išsivysto ant skirtingų individų, jie yra dvinamiai. Apvaisintas kiaušinis yra padengtas storu rudu lukštu; dažnai šalia jos esančios ląstelės suteikia trumpas šakeles, kurios auga virš oosporos, supindamos ją vienasluoksne žieve.
gyvavimo ciklai. Dauguma žaliųjų dumblių atstovų turi haplobionto gyvavimo ciklą su zigotiniu redukcija. Tokiose rūšyse tik zigota yra diploidinė stadija – ląstelė, susidaranti spermatozoidams apvaisinant kiaušinėlį. Kitas gyvenimo ciklo tipas – haplodiplobiontas su sporų mažinimu – aptinkamas Ulvoje, Cladophora ir kai kuriuose Trentepoliuose. Šie dumbliai pasižymi diploidinio sporofito ir haploidinio gametofito kaita. Haplodiplobionto gyvavimo ciklas su somatine redukcija žinomas tik Prasioloje. Kyla abejonių dėl diplobionto gyvavimo ciklo Briopsids ir Dasikladiis.
Kai kuriuose ulotriksuose tas pats individas gali sukelti zoosporas ir gametas. Kitais atvejais zoosporos ir gametos susidaro ant skirtingų individų, t.y. Dumblių gyvavimo ciklas apima ir seksualinę (gametofito), ir nelytinę (sporofito) vystymosi formas. Sporofitas dažniausiai yra diploidinis; ląstelėse turi dvigubą chromosomų rinkinį, gametofitas yra haploidinis, t.y. turi vieną chromosomų rinkinį. Tai pastebima tais atvejais, kai formuojantis sporoms (sporų redukcija) vyksta mejozė, o dalis dumblių gyvavimo ciklo nuo zigotos iki sporų susidarymo vyksta diplofazėje, o dalis nuo sporos iki gametų susidarymo. haplofazėje. Toks vystymosi ciklas būdingas Ulva genties rūšims.
Ulotrix dumbliuose zigotinė redukcija yra plačiai paplitusi, kai zigotai dygstant atsiranda mejozė. Tokiu atveju tik zigota pasirodo esanti diploidinė, o likusi gyvavimo ciklas vyksta haplofazėje. Daug rečiau pasitaiko gametinė redukcija, kai formuojantis gametoms atsiranda mejozė. Šiuo atveju tik gametos yra haploidinės, o likusi ciklo dalis – diploidinė.
Sistematika. Kol kas nėra vienos, nusistovėjusios žaliųjų dumblių sistemos, ypač kalbant apie užsakymų grupavimą į įvairias siūlomas klases. Labai ilgą laiką, skirstant žaliųjų dumblių užsakymus, pagrindinė reikšmė buvo teikiama talio diferenciacijos tipui. Tačiau pastaraisiais metais kaupiant duomenis apie žvynelinių ląstelių ultrastruktūrines ypatybes, mitozės ir citokinezės tipą ir kt., akivaizdus daugelio šių kategorijų nevienalytiškumas.
Skyriuje yra 5 klasės: Ulvophyceous – Ulvophyceae, Bripsodic – Bryopsidophyceae, Chlorophyceous – Chlorophyceae, Trebuxian – Trebouxiophyceae, Prasin – Prasinophyceae.
Ekologija ir prasmė.Žalieji dumbliai yra plačiai paplitę visame pasaulyje. Daugumą jų galima rasti gėlame vandenyje, tačiau yra daug sūrių ir jūrinių formų. Gijiniai žali dumbliai, prisitvirtinę arba laisvi, kartu su diatomėmis ir melsvai žaliomis dumbliais yra vyraujantys bentoso dumbliai žemyniniuose vandenyse. Jie randami skirtingo trofiškumo (nuo distrofinio iki eutrofinio) ir skirtingo organinių medžiagų (nuo kseno- iki polisaprobinių), vandenilio jonų (nuo šarminių iki rūgščių) vandens telkiniuose, esant skirtingoms temperatūroms (termo-, mezo- ir kriofilai). ).
Tarp žaliųjų dumblių yra planktoninių, perifitoninių ir bentosinių formų. Jūrinio pikoplanktono grupėje prazino dumbliai Ostreococcus yra laikomi mažiausia laisvai gyvenančia eukariotų ląstele. Yra žaliųjų dumblių rūšių, kurios prisitaikė gyventi dirvožemyje ir sausumos buveinėse. Jų galima rasti ant medžių žievės, uolų, įvairių pastatų, dirvos paviršiuje ir oro stulpelyje. Šiose buveinėse ypač paplitę Trentepolia ir Trebuxia genčių atstovai. Žalieji dumbliai vegetuoja karštuosiuose šaltiniuose, kurių temperatūra 35–52°C, o kai kuriais atvejais iki 84°C ir aukštesnė, dažnai su padidintu mineralinių druskų ar organinių medžiagų kiekiu (labai užterštos karštos nuotekos iš gamyklų, gamyklų, elektros energijos). elektrinės arba atominės elektrinės). Jie taip pat vyrauja tarp kriofilinių dumblių rūšių. Jie gali sukelti žalią, geltoną, mėlyną, raudoną, rudą, rudą ar juodą sniego ar ledo „žydėjimą“. Šie dumbliai randami paviršiniuose sniego ar ledo sluoksniuose ir intensyviai dauginasi apie 0 °C temperatūros tirpstančiame vandenyje. Tik kelios rūšys turi ramybės stadijas, o daugumai trūksta ypatingų morfologinių prisitaikymo prie žemos temperatūros.
Persūdytuose vandens telkiniuose vyrauja vienaląsčiai judrūs žalieji dumbliai - hiperhalobai, kurių ląstelėse nėra membranos ir jas supa tik plazmalema. Šie dumbliai išsiskiria dideliu natrio chlorido kiekiu protoplazmoje, dideliu intraląsteliniu osmosiniu slėgiu, karotinoidų ir glicerolio kaupimu ląstelėse, dideliu fermentų sistemų ir medžiagų apykaitos procesų labilumu. Sūrus vandens telkiniuose jų dažnai išsivysto daug, sukeldami raudoną arba žalią sūraus vandens telkinių „žydėjimą“.
Mikroskopinės vienaląstės, kolonijinės ir siūlinės žaliųjų dumblių formos prisitaikė prie nepalankių egzistavimo ore sąlygų. Priklausomai nuo drėgmės laipsnio, jie skirstomi į 2 grupes: oro dumbliai, gyvenantys tik atmosferos drėgmės sąlygomis, todėl nuolat kintantys ir džiūstantys; vandens dumbliai, nuolat drėkinami vandeniu (purškiant kriokliu, banglente ir pan.). Sąlygos dumbliams egzistuoti aerofilinėse bendruomenėse yra labai savotiškos ir joms pirmiausia būdingi dažni ir staigūs dviejų veiksnių – drėgmės ir temperatūros – pokyčiai.
Dirvos sluoksnyje gyvena šimtai žaliųjų dumblių rūšių. Dirvožemis kaip biotopas panašus ir į vandens, ir į oro buveines: jame yra oro, tačiau prisotintas vandens garų, o tai užtikrina atmosferos oro kvėpavimą be išdžiūvimo grėsmės. Intensyvus dumblių, kaip fototrofinių organizmų, vystymasis galimas tik šviesos prasiskverbimo ribose. Neapdorotose dirvose tai yra iki 1 cm storio paviršinis dirvožemio sluoksnis, dirbamose – šiek tiek storesnis. Tačiau dirvos storyje, kur šviesa neprasiskverbia, gyvybingi dumbliai aptinkami iki 2 m gylyje grynose dirvose ir iki 3 m gylyje ariamuose dirvožemiuose. Tai paaiškinama kai kurių dumblių gebėjimu tamsoje pereiti prie heterotrofinės mitybos. Daugelis dumblių lieka neaktyvūs dirvožemyje.
Norėdami išlaikyti savo gyvybinę veiklą, dirvožemio dumbliai turi tam tikrų morfologinių ir fiziologinių savybių. Tai yra palyginti nedideli dirvožemio rūšių dydžiai, taip pat gebėjimas gausiai formuotis gleives - gleivinės kolonijas, apvalkalus ir apvalkalus. Dėl gleivių, sudrėkinti dumbliai greitai sugeria vandenį ir jį kaupia, sulėtindami džiūvimą. būdingas bruožas dirvožemio dumbliai yra jų augmenijos „efemeriška prigimtis“ – gebėjimas greitai pereiti iš ramybės būsenos į aktyvų gyvenimą ir atvirkščiai. Jie taip pat gali toleruoti įvairius dirvožemio temperatūros svyravimus. Daugelio rūšių išgyvenimo diapazonas svyruoja nuo -200 iki +84 °C ir daugiau. Sausumos dumbliai sudaro svarbią Antarktidos augmenijos dalį. Jie nudažyti beveik juodai, todėl jų kūno temperatūra yra aukštesnė už temperatūrą aplinką. Dirvožemio dumbliai taip pat yra svarbūs sausringos (sausosios) zonos biocenozių komponentai, kur dirvožemis vasarą įšyla iki 60–80°C. Tamsūs gleiviniai dangteliai aplink ląsteles tarnauja kaip apsauga nuo per didelės insoliacijos.
Savotiškai grupei atstovauja endolitofiliniai dumbliai, susiję su kalkingu substratu. Pirma, tai yra dumblių gręžimas. Pavyzdžiui, Homontia genties dumbliai išgręžia miežių ir bedantukų kiautus ir gėlame vandenyje prasiskverbia į kalkingą substratą. Dėl jų kalkių substratas tampa purus, lengvai atsparus įvairiems cheminių ir fizinių veiksnių poveikiui. Antra, nemažai dumblių gėluose ir jūriniuose vandens telkiniuose sugeba vandenyje ištirpusias kalcio druskas paversti netirpiomis ir nusodinti jas ant savo talio. Daugelis atogrąžų žaliųjų dumblių, ypač Galimeda, kaupia kalcio karbonatą taluose. Jie priima Aktyvus dalyvavimas rifo pastate. Milžiniškos Galimedos liekanų nuosėdos, kartais siekiančios 50 m aukštį, aptinkamos žemyninio šelfo vandenyse, susijusiuose su Didžiuoju barjeriniu rifu Australijoje ir kituose regionuose, 12–100 m gylyje.
Žalieji trebukso dumbliai, užmezgantys simbiotinį ryšį su grybais, yra kerpių dalis. Apie 85% kerpių yra vienaląsčių ir siūlinių žaliųjų dumblių kaip fotobiontų, 10% - melsvadumblių, o 4% (ar daugiau) yra ir melsvadumblių, ir žaliųjų dumblių. Jie egzistuoja kaip endosimbiontai pirmuonių, kriptofitų, hidrų, kempinių ir kai kurių plokščiųjų kirmėlių ląstelėse. Net atskirų sifoninių dumblių, tokių kaip kodis, chloroplastai tampa nudibranch moliuskų simbiontais. Šie gyvūnai minta dumbliais, kurių chloroplastai išlieka gyvybingi kvėpavimo ertmės ląstelėse, o šviesoje labai efektyviai fotosintezuoja. Ant žinduolių kailio išsivysto nemažai žaliųjų dumblių. Endosimbiontai, patiriantys morfologinius pokyčius, palyginti su laisvai gyvenančiais atstovais, nepraranda gebėjimo fotosintezuoti ir daugintis šeimininko ląstelėse.
Ekonominė vertė. Dėl savo gebėjimo fotosintezuoti jie yra pagrindiniai didžiulio kiekio organinių medžiagų gamintojai vandens telkiniuose, kuriuos plačiai naudoja gyvūnai ir žmonės. Sugerdami anglies dioksidą iš vandens, žalieji dumbliai prisotina jį deguonimi, reikalingu visiems gyviems organizmams. Puikūs jie vaidmenį biologiniame medžiagų cikle. Greitas dauginimasis ir labai didelis asimiliacijos greitis (maždaug 3-5 kartus didesnis nei sausumos augalų) lemia tai, kad dumblių masė per dieną padidėja daugiau nei 10 kartų. Tuo pačiu metu chlorelės ląstelėse kaupiasi angliavandeniai (veislinėse padermėse jų kiekis siekia 60 proc.), lipidai (iki 85 proc.), vitaminai B, C ir K. Chlorella baltymai gali sudaryti iki 50 proc. sausos ląstelės masės, yra visų nepakeičiamų aminorūgščių. Unikalus Chlorella rūšių gebėjimas pasisavinti nuo 10 iki 18 % šviesos energijos (palyginti su 1–2 % sausumos augalų) leidžia panaudoti šiuos žaliuosius dumblius oro regeneracijai uždarose biologinėse žmogaus gyvybės palaikymo sistemose ilgalaikių skrydžių metu kosmose. ir nardymas.
Nemažai žaliųjų dumblių rūšių yra naudojami kaip indikatoriniai organizmai vandens ekosistemų monitoringo sistemoje. Kartu su fototrofiniu maitinimosi būdu daugelis vienaląsčių žaliųjų dumblių (chlamidomonų) sugeba per kiautą pasisavinti vandenyje ištirpusias organines medžiagas, o tai prisideda prie aktyvaus užteršto vandens, kuriame vystosi šios rūšys, valymo. Todėl jie naudojami užterštų vandenų, taip pat pašarų žuvininkystės rezervuaruose valymui ir tolesniam valymui.
Kai kurių rūšių žaliuosius dumblius daugelio šalių gyventojai naudoja maistui.
Paviršinės žaliųjų dumblių plėvelės turi didelę antierozinę vertę. Ląstelių membranų gleivinės sulipina dirvožemio daleles. Dumblių vystymasis daro įtaką smulkios žemės struktūrai, todėl ji yra atspari vandeniui ir neleidžia jai nunešti nuo paviršinio sluoksnio.
Dirvožemio dumbliai taip pat turi įtakos aukštesnių augalų augimui ir vystymuisi. Išskirdami fiziologiškai aktyvias medžiagas, jie pagreitina daigų, ypač jų šaknų, augimą.
Dumblių ląstelės sugeba iš vandens kaupti įvairius cheminius elementus, o jų kaupimosi koeficientai gana dideli. Galingi koncentratoriai yra gėlavandeniai žalieji dumbliai, ypač siūliniai. Tuo pačiu metu metalų kaupimosi intensyvumas juose yra daug didesnis nei kituose gėlavandeniuose hidrobiontuose. Didelį susidomėjimą kelia dumblių gebėjimas koncentruoti radioaktyvius elementus. Negyvos dumblių ląstelės sukauptus elementus išlaiko ne mažiau tvirtai nei gyvosios, o kai kuriais atvejais desorbcija iš negyvų ląstelių yra mažesnė nei iš gyvųjų. Daugelio genčių (Chlorella, Scenedesmus ir kt.) gebėjimas sutelkti ir tvirtai išlaikyti savo ląstelėse cheminius elementus ir radionuklidus leidžia juos naudoti specializuotose valymo sistemose, skirtose pramoninėms nuotekoms nukenksminti, pavyzdžiui, papildomam valymui. žemo aktyvumo nuotekų iš atominių elektrinių.
Kai kurie žalieji dumbliai yra gripo viruso, polioviruso ir kt. antagonistai. Dumblių išskiriamos biologiškai aktyvios medžiagos atlieka svarbų vaidmenį dezinfekuojant vandenį ir slopinant patogeninės mikrofloros gyvybinę veiklą.
Specialiuose biologiniuose tvenkiniuose dumblių ir bakterijų bendrijos naudojamos herbicidams skaidyti ir detoksikuoti. Įrodytas daugelio žaliųjų dumblių gebėjimas hidrolizuoti herbicidą propanilį, kurį greitai sunaikina bakterijos.
Aukštesniųjų augalų audinių diferenciacija, jos atsiradimas evoliucijoje. Audinių rūšys.
Tarp Žemėje gyvenančių augalų yra vienaląsčių, kurių kūnas susideda iš vienos ląstelės. Tai chlorelės dumbliai, chlamidomonas. Šiuose augaluose ląstelė yra atskiras holistinis organizmas. Jame vyksta visi gyvybės procesai: mityba, kvėpavimas, medžiagų susidarymas ir išsiskyrimas, dauginimasis.
Augalai, kurių kūną sudaro daug ląstelių, vadinami daugialąsčiais. Dauguma šių augalų. Tai kai kurie dumbliai, samanos, paparčiai, gimnasėkliai ir žydintys augalai. Daugialąsčiuose augaluose ląstelės dažniausiai skiriasi struktūra ir funkcija. Vieni atlieka mitybos, kiti – dauginimosi, treti – medžiagų judėjimo funkciją.
Augalų struktūra istorinės raidos procese pamažu komplikavosi, daugėjo įvairių tipų ląstelių. Jei vienaląsčiuose dumbliuose kūnas susideda iš vienos ląstelės, tai samanose jau yra apie 20 skirtingų tipų ląstelių, paparčiuose - apie 40, o gaubtasėkliuose - apie 80.
Ląstelių grupės, kurios yra panašios struktūros, kilmės ir pritaikytos atlikti vieną ar daugiau funkcijų, vadinamos audiniu. Audiniai, susidedantys iš vieno tipo ląstelių, vadinami paprastais, o tie, kurie susideda iš skirtingų tipų, vadinami kompleksiniais arba kompleksiniais.
Augaluose yra lavinamieji, vientisi, mechaniniai, laidūs, fotosintezės ir saugojimo audiniai.
Ugdomasis audinys (meristema). Šis audinys susideda iš daugiau ar mažiau identiškų ląstelių su plonomis membranomis, galinčiomis dalytis. Ląstelės tvirtai priglunda viena prie kitos, turi branduolį, citoplazmą, neturi pastebimų vakuolių. Jie išsidėstę šaknų ir ūglių galiukuose, jaunų lapų apačioje, stiebų tarpubambiuose, po medžių kamienų žieve (kambis). Ugdomojo audinio ląstelės nuolat dalijasi, dėl to auga ūglis ir šaknis į ilgį ir storį, žydi pumpurai, pumpurai, iš sėklų išauga daigai. Ugdomasis audinys užtikrina augalų augimą, naujų audinių ir organų formavimąsi.
Dangos audinys. Šio audinio ląstelės tvirtai prilimpa viena prie kitos ir saugo augalo organus nuo neigiamo išorinės aplinkos poveikio: džiūvimo, mechaninių pažeidimų. Integruoto audinio dėka augalas sąveikauja su aplinka, per jį į ląstelę patenka reikalingos medžiagos iš aplinkos ir išsiskiria atliekos. Pavyzdžiui, per augalo lapo vientisą audinį (stomatas) vyksta dujų mainai ir vanduo išgaruoja.
mechaninis audinys. Šį audinį sudaro ląstelės su storomis, dažnai labai stipriomis membranomis, kurios dažnai yra impregnuotos į riebalus panašiomis medžiagomis. Todėl audinys suteikia augalui nuolatinę formą, užtikrina jo atsparumą lūžimui ir lenkimui. Mechaniniai audiniai sudaro augalo skeletą ir suteikia jam tvirtumo, todėl jie dar vadinami atraminiais audiniais.
Fotosintetinis (asimiliacinis) audinys. Šis audinys susideda iš plonasienių gyvų ląstelių, kurių citoplazmoje yra daug chloroplastų. Jie fotosintezės metu gamina organines medžiagas. Fotosintetinis audinys yra žalias. Taip yra dėl to, kad chloroplastuose yra chlorofilo – žalio pigmento. Fotosintetinio audinio ląstelėse, be chlorofilo, yra ir nedideli kiekiai kitų pigmentų (geltonų, oranžinių). Jie sugeria trumpųjų bangų spindulius ir veikia kaip ekranas, apsaugantis chlorofilą ir ląstelę nuo žalingo šių spindulių poveikio.
laikymo audinys. Jį sudaro didelės gyvos ląstelės su plonomis membranomis. Juose nusėda atsarginės medžiagos: baltymai, riebalai ir angliavandeniai. Vienuose augaluose atsarginės medžiagos nusėda į sėklas, kitų – į šaknis, stiebus, gumbus ir kt. Kaktusų stiebuose, alavijo lapuose gerai išvystytas vandenį sulaikantis audinys, leidžiantis augalams atlaikyti saulės šilumą, taip pat vandens trūkumą dirvoje. Vartodami vandenį, augalų audiniai praranda savo elastingumą, tačiau įsisavinę vandenį atstato ankstesnis vaizdas ir dydžiai.
Laidūs audiniai prasiskverbia per visą augalo kūną, sudarydami ištisinę šakotą sistemą – nuo mažiausių šaknų iki jauniausių lapelių. Laidieji audiniai prisideda prie kylančios ir besileidžiančios srovės susidarymo augale. Kylanti srovė – tai vandenyje ištirpusių mineralinių druskų srovė, einanti nuo šaknų išilgai stiebo iki lapų. Kylanti srovė teka per ksilemo indus (trachėjus) ir tracheidus. Žemyn nukreipta srovė yra organinių medžiagų srovė, nukreipta iš lapų į šaknis išilgai floemo sieto elementų. Seniausi laidūs ksilemo elementai yra tracheidės – pailgos ląstelės smailiais galais. Tracheidų ląstelės turi lignifikuotą ląstelių sienelę, kurios sustorėjimas yra įvairus. Yra žiedinės, spiralinės, taškinės, porėtos ir kitos tracheidės. Indai yra ilgi tuščiaviduriai vamzdeliai, sudaryti iš vienos eilės negyvų ląstelių, kurių skersinės pertvaros ištirpo. Floemo sieto vamzdeliai yra gyvos ląstelės, pailgos, kurių galuose (ląstelių sąlyčio taškuose) yra sieto skylutės. Floemas ir ksilemas sudaro kraujagyslių pluoštinius ryšulius. Floemas ir ksilemas yra sudėtingi audiniai. Taigi, ksilemas apima kraujagysles, tracheidas, parenchimą ir medienos pluoštus (ne visada). Floemą sudaro sieto vamzdeliai ir kompanioninės ląstelės, parenchima ir kartais karnienos pluoštai.
Aukštųjų augalų dauginimosi rūšys.
I. Dauginimasis, lydimas dauginimosi: A - motinos individas, A - dukra, panaši į motiną.
II. Dauginimasis, be dauginimosi: W - motininis individas, nustoja egzistuoti susiformavus dukrai.
III. Dauginimas be dauginimo. Motina A susilaukia palikuonių B, kurie nėra panašūs į ją.
IV. Palikuonių, kurie nėra dauginimasis (B palikuonis nėra panašus į motiną A), auginimas ir nėra lydimas dauginimosi.
Augalų dauginimo rūšys. Nauji individai gali būti formuojami iš tėvų nelytiškai, tai yra nedalyvaujant lytinėms ląstelėms ir lytiniam procesui, ir lytiškai, kai prieš dukterinių individų formavimąsi būtinai vyksta lytinis procesas. Nelytinio dauginimosi plačiąja prasme atveju motininis individas arba dalijasi į daugiau ar mažiau lygias dalis (vienaląsčių dumblių dalijimasis, dalijimasis daugiamečių žolių, žr. V ir VII skyrius), arba atskiria nuo savęs mažus dukterinių individų, galinčių išsivystyti į pradmenis. nepriklausomi augalai. Vegetatyvinio kūno dalijimasis arba vegetatyvinių užuomazgų, pavyzdžiui, pumpurų, mazgelių, atskyrimas nuo jo yra vegetatyvinis nelytinis dauginimasis. Tuo pačiu metu palikuonių genotipas išlieka iš esmės nepakitęs ir kiekvienas palikuonis "visiškai atgamina motininį augalą. Kartais specializuotos ląstelės, vadinamos sporomis, yra atskirtos kaip rudimentai (gr. sporas - sėja, sėja). Aukštesniuose augaluose sporų susidarymas yra susijęs su chromosomų skaičiaus sumažėjimu, todėl iš sporų augantys dukteriniai augalai nepanašūs į motininius augalus (jų nesidaugina).Nors sporų dauginimasis yra nelytinis, kadangi sporos dygsta nesusiliejusios su kitomis ląstelėmis, yra susijusios. aukštesniuose augaluose, kurių lytinis dauginimasis vyksta reguliariu dauginimosi ciklu (žr. toliau) .
Lytinio dauginimosi metu dukterinių individų genotipas gali keistis ir turtėti dėl įvairių tėvų individualių savybių rekombinacijų.
Bendrosios vegetatyvinio dauginimo charakteristikos. Vegetatyvinis dauginimas – tai tam tikros rūšies ar veislės individų skaičiaus didinimas, atskiriant gyvybingas augalo vegetatyvinio kūno dalis. Kiekviena atskirta dalis kurį laiką gyvena savarankiškai ir, kaip taisyklė, suformuoja naujus organus, kurių dažnai trūksta (atskirtame ūglyje formuojasi šaknys, ant šaknies dalių formuojasi ūgliai). Taigi vegetatyvinio dauginimo metu įprastas ir būdingas yra regeneravimas - visumos atstatymas iš dalies. Tačiau gana dažnai būsimam savarankiškam individui visi reikalingi organai susikuria dar jam neatskiriant nuo motinos (pavyzdžiui, nauji rozetiniai ūgliai su papildomomis šaknimis braškių ūsų galuose).
Briofitų ir paparčių vystymosi ciklas, susijęs su prisitaikymu prie sausumos gyvenimo sąlygų.
Paparčiai turi būdingą, neprilygstamą išvaizdą. Tai sumedėjęs arba žolinis augalas. Turi modifikuotą ūglį, prie kurio lapkočio pagalba pritvirtinami pseudolapai, arba gniuželiai. Tai pirmasis evoliucinis žingsnis, siekiant suformuoti tikrą augalų lapų geležtę. Vayi atlieka dvi funkcijas: pirmoji yra fotosintezė, antroji - sporuliacija.
Žemėje augalas tvirtinamas požeminio stiebo – šakniastiebio pagalba. Daugelis vegetatyvinių šaknų išsiskiria iš jo. Paparčio stiebe susidaro audiniai - laidūs ir parenchiminiai, suteikiantys augalui galimybę suvartoti daugiau mineralų ir vandens nei jo žemesnio organizacinio atitikmenys planetoje.
Paparčio gyvenimo ciklas susideda iš dviejų fazių - sporofito ir gametofito, o pirmoji fazė vyrauja prieš antrąją. Apatinėje lapelio dalyje susidaro haploidinės sporos. Laikui bėgant sporos atsidaro, sporos nukrenta ant žemės ir sudygsta. Būtent šis augimas perneša moteriškas ir vyriškas lytines ląsteles. Tačiau to paties augalo kiaušinėliai ir spermatozoidai subręsta skirtingu laiku, todėl savaiminis apvaisinimas nevyksta. Paparčiams, kaip ir samanoms, klestėti reikalinga labai drėgna aplinka.
Apvaisintas zigotas virsta sporofitu. Iš pradžių ji naudoja maistines medžiagas, kurios yra augime, o kai jis miršta, pradeda maitintis pati.
Samanos yra dvinamiai augalai, tai yra, vyriškų augalų viršūnėse yra organai, gaminantys spermatozoidus, o moteriškų augalų viršūnėse yra kiaušinėlių gamintojai. Bet kiekvienas augalas, nepriklausomai nuo lyties, turi stiebą ir lapus. Jie yra maži, juose yra chlorofilo. Daugelyje samanų apatinės pakopos lapai tampa geltonai rudi dėl pigmento sunaikinimo esant prastam apšvietimui.
Samanos neturi šaknų. Prie žemės juos pritvirtina rizoidai – daugialąsčiai į plauką panašūs procesai.
Samanos dauginasi sporomis, kurios subręsta sporofito sporangijose. Samanų sporofitą vaizduoja koja su dėžute. Bet jis ilgai negyvena ir greitai išdžiūsta. Išdžiūvusi dėžutė atsidaro, ir iš jos pabunda sporos. Iš jų išauga augalas su haploidiniu chromosomų rinkiniu – daugiametis, žalias; moteris ar vyras. Samanų gyvavimo cikle dominuoja gametofitas, o ne sporofitas.
Moteriškojo ir vyriškojo gametofito sandara ir vystymasis gimnasėkliuose ir gaubtasėkliuose.
Moteriškas gametofitas visuose gimnosėkliuose visiškai išsivysto megasporangiumo viduje ir net iš dalies neišeina į lauką, tai yra, tiesiogiai nesiliečia su oru. Į moterišką gametofitą patenkama tik per mikropilą. Taigi kiaušialąstės viduje susidaro palankiausios sąlygos apsaugoti moterišką gametofitą nuo išdžiūvimo. Dėl to pamažu mažėja ir supaprastėja patelės gametofitas ir archegonija, atsiranda galimybė labai anksti susiformuoti kiaušinėliui, o kai kuriuose gimnosėkliuose (velvichia ir gnetum) susidaro net specialūs neoteniniai nearchegoniniai gametofitai.
Gimnosėkliai nuo paparčių skiriasi ir vyriškojo gametofito išsivystymu, mikrosporų sandara ir dygimo būdu. Paparčiuose, kur gametofitas vystosi dažniausiai tik pasėjus sporas, sporos dygsta per vadinamąjį tetradinį randą, esantį proksimaliniame sporos poliuje. Gimnosėkliuose, kur vyriškasis gametofitas yra labai supaprastintas ir pagreitėja jo vystymasis, pirmieji mikrosporos branduolio pasiskirstymai vyksta jau mikrosporangijos viduje. Atsižvelgiant į ankstyvą vyriškojo gametofito vystymąsi ir lytinių ląstelių susidarymą net sporų membranos viduje, reikia prisitaikyti, kad mikrosporos galėtų pakeisti savo tūrį. Toks prisitaikymas yra mikrosporos distaliniame poliuje esanti vaga, kuri pirmiausia atsiranda kai kuriuose sėkliniuose paparčiuose ir būdinga didžiajai daugumai gimnasėklių. Vaga tarnauja ne tik žiedadulkių grūdų tūriui reguliuoti. Tai tampa išėjimo iš haustorijos mikrosporų (žemesnėse grupėse) arba žiedadulkių vamzdelio (spygliuočių ir spygliuočių), kurie taip pat yra navikai, vieta. Taigi gimnosėkliuose, priešingai nei paparčiuose, anga mikrosporos turiniui išleisti susidaro distaliniame poliuje. Cikadinio tipo gaustoria (siurblys) auga horizontaliai ir skirta tik pritvirtinti ir maitinti vyrišką gametofitą; tikrasis spygliuočių ir gneataceae žiedadulkių vamzdelis auga vertikaliai ir daugiausia naudojamas spermatozoidams pernešti į kiaušinėlius, tai yra, jis yra laidininkas (vektorius), o ne tik siurblys. Nors abu šie dariniai paprastai vadinami žiedadulkių vamzdeliais, morfologiškai ir funkciškai jie labai skiriasi.
Galiausiai reikia pažymėti, kad zigotos branduolinis (branduolinis) padalijimas būdingas gimnasėkliams (išskyrus velvichijas, gnetum ir visžalius sekvojus). Šiuo požiūriu jie skiriasi ne tik nuo žemesnių grupių, bet ir nuo gaubtasėklių, kuriems (išskyrus tik pion gentį) būdingas zigotos ląstelinis suskaidymas.
Gimnosėklių vystymosi ciklai yra tarsi pereinamasis etapas tarp paparčių ir gaubtasėklių vystymosi ciklų. Taigi, pavyzdžiui, pušis sudaro dviejų rūšių kūgius - mažus patinus, kurių ilgis ne didesnis kaip 2,5 cm, ir didelę patelę, kai kurių rūšių ilgio siekia 45 cm. Moteriškas kūgis susideda iš daugybės žvynų, kurių kiekvieno paviršiuje yra dvi kiaušialąstės. Kiekvienoje kiaušialąstėje yra diploidinė makrosporos motininė ląstelė. Pastaroji dalijasi mejotiškai, susidarant keturioms haploidinėms makrosporoms, iš kurių funkcionuoja tik viena ir išsivysto daugialąstelinis makrogametofitas. Ant kiekvieno tokio makrogametofito yra 2–3 moteriški lytiniai organai (archegonija), kuriuose yra vienas didelis kiaušinis. Kiekvienos vyriškojo kūgio žvyno apačioje yra dvi mikrosporangijos. Šiose mikrosporangijose yra daug mikrosporinių motininių ląstelių, kurių kiekviena dalijasi mejotiškai ir sudaro keturias mikrosporas. Mikrosporangiume arba žiedadulkių maišelyje esančios mikrosporos dalijasi ir sudaro keturių ląstelių mikrogametofitą arba žiedadulkių grūdus. Laisvas žiedadulkes neša vėjas. Pasiekę patelės kūgį, žiedadulkės pro specialią angą – mikropilį – prasiskverbia į kiaušialąstę ir susiliečia su makrosporangiu.
Gali praeiti daugiau nei metus prieš tai, kai viena iš žiedadulkių grūdelių ląstelių išsivysto į žiedadulkių vamzdelį, kuris išauga per makrosporangumą ir pasiekia makrogametofitą. Dar viena žiedadulkės ląstelė dalijasi, susidarant ne judriam spermatozoidui, kaip žemesniuose augaluose, o du vyriškus generatyvinius branduolius. Kai žiedadulkių vamzdelio galas pasiekia archegonijos kaklelį ir atsidaro, iš jo išeina du vyriški branduoliai, esantys šalia kiaušinėlio. Vienas iš jų susilieja su kiaušinėlio branduoliu, sudarydamas diploidinę zigotą, o kitas išnyksta. Po apvaisinimo zigota dalijasi ir diferencijuojasi, suformuodama sporofito embrioną, apsuptą moteriškojo gametofito audinių, taip pat motininio sporofito audinių. Visas šis kompleksas yra sėkla.
Makrogametofito audiniai, aprūpinantys besivystantį embrioną maistinėmis medžiagomis, sudaro endospermą. Tačiau jas sudaro haploidinės ląstelės, o ne triploidinės (3n) ląstelės, tokios kaip angiospermo endospermo ląstelės, nors abi yra skirtos embriono mitybai. Po trumpo augimo laikotarpio, per kurį susiformuoja keli lapo formos skilčialapiai, epikotilas (sukeliantis stiebą) ir hipokotilas (sukeliantis pirmines šaknis), embrionas pereina į ramybės būseną ir išlieka tokioje būsenoje, kol nukrenta. į žemę. Patekusi į palankias sąlygas, sėkla sudygsta ir išsivysto subrendęs sporofitas – pušis.
Gaubtasėklių vystymosi ciklas.
Gaubtasėkliuose vis dar vyksta kartų kaita – sporofitas ir gametofitas, tačiau gametofitas redukuojasi iki kelių ląstelių, esančių sporofito žiedo audiniuose. Sporofitai – tai paprasti mums gerai žinomi medžiai, krūmai ar žolelės. Ne visų augalų žiedai yra lengvai atskiriami; maži žali javų ir kai kurių medžių žiedai gerokai skiriasi nuo tų ryškiaspalvių darinių, kuriuos paprastai vadiname gėlėmis.
Gaubtasėklio gėlė yra modifikuotas ūglis, kuriame vietoj įprastų žalių lapų yra koncentriškai išsidėstę lapai, modifikuoti taip, kad atliktų dauginimosi funkciją. Įprastą gėlę sudaro koncentriškai išdėstyti keturių tipų elementai, pritvirtinti prie talpyklos - išplėstinio žydinčio stiebo galo. Tolimiausi elementai – taurėlapiai – dažniausiai yra žali ir labiausiai panašūs į tikrus lapus. Taurėlapių žiede yra žiedlapių, kurių dauguma yra ryškios spalvos, kad pritrauktų vabzdžius ar paukščius apdulkinti.
Tiesiai žiedlapių žiedo viduje yra kuokeliai – vyriškosios gėlės dalys. Kiekvienas kuokelis susideda iš plonos gijos, kurios gale yra dulkinys. Dulkinė yra žiedadulkių maišelių (mikroporangijų) grupė, kurių kiekvienoje yra mikrosporinių motininių ląstelių, vadinamųjų žiedadulkių motininių ląstelių. Dėl mejozės kiekviena iš šių diploidinių ląstelių sudaro keturias haploidines mikrosporas, kurios po branduolio dalijimosi virsta jaunais mikrogametofitais arba žiedadulkių grūdeliais.
Pačiame gėlės centre yra piestelių žiedas (arba viena piestelė, susidariusi susiliejus kelioms piestelėmis). Piestelė susideda iš sustorėjusios tuščiavidurės apatinės dalies – kiaušidės ir iš jos besitęsiančios ilgos plonos kolonos, kuri viršuje baigiasi išlyginta stigma. Pastarasis dažniausiai išskiria lipnų skystį, kuris sulaiko ir sulaiko ant grūstuvės nukritusias žiedadulkes. Visos šios gėlės dalys yra labai įvairios savo skaičiumi, išdėstymu ir forma. Gėlė, kurioje yra ir kuokelių, ir piestelių, vadinama biseksualia; gėlės be kuokelių ar piestelių yra vienalytės. Tos pačios lyties gėlės, turinčios tik kuokelius, vadinamos kuokelėmis; vienalytės gėlės, kuriose yra tik piestelės, vadinamos piestelėmis. Gluosniai, tuopos ir datulinė palmė yra tarp augalų, kurių vieni žiedai neša tik kuoduotus žiedus, o kiti – tik piestelinius. Žydinčių augalų dauginimosi organai – kuokeliai, piestelės, stigma, stilius ir kt. – buvo tiriami ir pavadinti dar prieš išaiškinant įvairiems kartų kaitos etapams ir iki pagrindinių samanų vystymosi ciklo ypatybių lygiagretumo, t. buvo atskleisti paparčiai ir žydintys augalai.
Kiaušidėje, esančioje piestelės apačioje, yra viena ar daugiau kiaušialąsčių. Pastarieji yra makrosporangiai, apsupti 1 arba 2 sluoksniais. Paprastai kiekvienoje kiaušialąstėje yra viena makrosporos motininė ląstelė, kuri dėl mejozės sudaro keturias haploidines makrosporas. Viena iš makrosporų išsivysto į makrogametofitu; kiti trys sunaikinti. Makrogametofito vystymosi eiga kiekvienai rūšiai yra specifinė; tipiniu atveju makrospora labai padidėja ir jos branduolys dalijasi. Du dukteriniai branduoliai migruoja į priešingus ląstelės galus, kiekvienas iš jų dalijasi, o tada šie dukteriniai branduoliai taip pat dalijasi. Taip susidaręs makrogametofitas, vadinamas embriono maišeliu, yra aštuonių branduolių ląstelė, kurios kiekviename gale yra keturi branduoliai. Vienas branduolys iš kiekvieno galo juda į ląstelės centrą; šie du vienas šalia kito esantys ląstelės centre esantys branduoliai vadinami poliniais branduoliais. Vienas iš trijų viename makrosporofito gale esančių branduolių tampa kiaušinėlio branduoliu, o kiti du ir trys kitame gale esantys branduoliai išnyksta.
Haploidinė mikrospora žiedadulkių maišelio viduje išsivysto į mikrogametofitą arba žiedadulkių grūdus. Mikrosporos branduolys dalijasi, sudarydamas didelį žiedadulkių vamzdelio branduolį ir mažesnį generacinį branduolį. Daugeliu atvejų žiedadulkės išsiskiria šiame etape ir vėjo, vabzdžių ar paukščių pernešamos ant tos pačios arba šalia esančios gėlės stigmos. Patekusios į stigmą, žiedadulkės sudygsta. Kai žiedadulkės sudygsta, susidaro žiedadulkių vamzdelis, augantis stulpeliu iki kiaušialąstės. Žiedadulkių vamzdelio galas išskiria fermentus, kurie ištirpdo stiliaus ląsteles, kad būtų galima toliau dygti. Vamzdžio šerdis lieka augančio žiedadulkių vamzdelio gale. Generatyvinis branduolys migruoja į žiedadulkių vamzdelį ir dalijasi suformuodamas du branduolius – spermatozoidų branduolius. Subrendęs vyriškas gametofitas susideda iš žiedadulkių grūdelių ir žiedadulkių vamzdelio, vamzdelio branduolio ir dviejų spermatozoidų branduolių bei kai kurios susijusios citoplazmos.
Per mikropilą prasiskverbę į makrogametofitą, žiedadulkių vamzdelio galiukas plyšta ir abu generatyviniai branduoliai prasiskverbia į makrogametofitą. Vienas iš šių branduolių persikelia į kiaušinėlio branduolį ir susilieja su juo; susidaręs diploidinis zigotas sukelia naujos kartos sporofitus. Kitas generacinis branduolys pereina į du poliarinius branduolius, po kurių visi trys branduoliai susilieja ir sudaro endospermo branduolį, kuriame yra trigubas chromosomų rinkinys. Kartais du poliariniai branduoliai susilieja į vieną dar prieš atsirandant generatyviniam branduoliui. Aprašytas dvigubo apvaisinimo reiškinys, lemiantis diploidinio zigoto ir triploidinio (su trigubu chromosomų rinkiniu) endospermo atsiradimą, yra specifinis ir būdingas žydintiems augalams.
Po apvaisinimo zigota daug kartų dalijasi ir suformuoja daugialąstį embrioną. Dėl endospermo branduolio dalijimosi susidaro endospermo ląstelės, užpildytos maistinėmis medžiagomis. Šios ląstelės, supančios embrioną, aprūpina jį maistu. Po apvaisinimo taurėlapiai, žiedlapiai, kuokeliai, stigma ir stilius dažniausiai nuvysta ir nukrinta. Kiaušialąstė kartu su joje esančiu embrionu virsta sėkla; jo sienelės sustorėja ir virsta kietais išoriniais sėklos gaubtais. Sėkla susideda iš embriono ir endospermo su maistinėmis medžiagomis, uždengtais tvirtame lukšte, kuris atsirado iš kiaušialąstės sienelės. Sėklų dėka rūšis turi galimybę plisti į naujas buveines ir išgyventi nepalankių išorinių sąlygų periodus (pavyzdžiui, žiemą), kurios kenkia suaugusiems augalams.
Vaisius. Kiaušidės – apatinė piestelės dalis, kurioje yra kiaušialąstės – auga ir virsta vaisiumi. Taigi vaisiuose esančių sėklų skaičius atitinka kiaušialąsčių skaičių. Griežtai botanine šio žodžio prasme vaisius yra subrendusios kiaušidės, kuriose yra sėklos – subrendusios kiaušialąstės. Kasdieniame gyvenime vaisiais vadiname tokius kvapnius, mėsingus darinius kaip vynuogės, uogos, obuoliai, persikai, vyšnios. Tačiau pupelės ir žirniai, kukurūzų branduoliai, pomidorai, agurkai ir melionai, taip pat riešutai, varnalėšos ir sparnuoti klevai taip pat yra vaisiai. Tikrasis vaisius išsivysto tik iš kiaušidės. Vaisiai, atsirandantys iš taurėlapių, žiedlapių ar talpyklos, vadinami netikru vaisiumi. Obuolių vaisius daugiausia sudaro peraugęs mėsingas indas; tik obuolio šerdis ateina iš kiaušidės.
Vegetatyvinių organų metamorfozė.
Metamorfozės – tai organų modifikacijos, atsiradusios augalų evoliucijos procese pasikeitus ar pasikeitus organų funkcijoms, veikiant aplinkos sąlygų kompleksui.
Šakniavaisiai ir šakniavaisiai skirti kaupti medžiagas, reikalingas sėkloms formuotis. Šakniavaisiai susiformuoja, kai sustorėja pagrindinė šaknis ir apatinė ūglio dalis (burokėliai, rūtos, ropės, ropės, ridikai, ridikai, morkos, petražolės, stulpeliai, kiaulės), o ant šoninių šaknų (jurginai šparagai, gumbinis agrastas, šešiažiedis pievinis smėlis) dvimečiai arba daugiamečiai augalai.
Atsitraukiančios šaknys gali sutrumpėti prie jų pagrindo. Dėl to jie giliau į dirvą traukia svogūnėlius, šakniastiebius, gumbus, kur jų inkstai yra apsaugoti nuo nepalankių veiksnių. Atsitraukiančias šaknis nesunku atpažinti iš skersinių juostelių ant sustorėjusių pagrindų. Jie būdingi tokiam augalui kaip lelija - saranka, kurioje su amžiumi svogūnėlis vis giliau patenka į dirvą. Gerai išreikštas kardelių gumbasvogūniais. Akivaizdu, kad jų galima rasti kituose augaluose su svogūnėliais, šaknų spurgais ir skęstančiomis šakniastiebiais. Yra žinoma, kad žemuogėse, plaučiuose, kanopose, žibuoklėse, rankogalyje, gravilata, antžeminiai ūgliai su žvynuotais ir žaliais rozetės lapais praranda fotosintetinius lapus ir yra įtraukiami į dirvą atsitiktinių šaknų, tampa šakniastiebiais.
bakteriniai mazgeliai. Daugelio augalų šoninės šaknys yra pritaikytos simbiozei su tam tikromis azotą fiksuojančiomis bakterijomis. Iš molekulinio oro azoto mazgeliuose sintetinamos organinės medžiagos, kurių dalį augalas panaudoja. Be ankštinių augalų, jie sugeba suformuoti mazgelius ant snukių, alksnio, šaltalankių šaknų. Ankštinėse daržovėse dėl papildomo azoto šaltinio gausu baltymų. Jie suteikia vertingų maisto ir pašarų produktų, praturtina dirvą azotinėmis medžiagomis (200-300 kg azoto 1 hektarui dirvožemio), todėl dažnai naudojami kaip „žaliosios trąšos“, o šaltalankiai – melioruojant pažeistas žemes.
mikorizė (grybelio šaknis). Daugelio augalų šaknys gyvena kartu su dirvožemio grybais, sudarydamos mikorizę („grybelio šaknį“). Grybeliniai hifai padeda šaknims lengviau pasisavinti vandenį ir mineralines medžiagas iš dirvos, perneša jas tam tikroms organinėms medžiagoms, vitaminams, augimo reguliatoriams. Grybelis angliavandenius ir kitas maistines medžiagas gauna iš aukštesnių augalų. Susiformuoja mikorizė dauguma laukinės ir kultūrinės žolės ir medžiai - gimnazdžių, vienaląsčių (75%) ir dviskilčių (80-90% viso rūšių skaičiaus) atstovai. Augalai, pavyzdžiui, sumedėję augalai, geriau auga esant grybeliui, o orchidėjų sėklos sudygsta tik tokiu atveju. Medžiuose grybų gijos dažniausiai yra išorėje (apvalkalo pavidalu) ir šaknų paviršiniuose audiniuose, kuriuose šiuo atveju nėra šaknų plaukų. Kai gyvena kartu su žolelėmis, grybų sporos paprastai prasiskverbia į šaknų vidų, kurios gali intensyviai šakotis ir formuoti patinimus. Rūgštus lietus sukelia mikorizę formuojančių grybų rūšinės įvairovės sumažėjimą ir mikorio sunaikinimą.Šie duomenys gauti tiriant pušį, beržą ir eglę! Escape modifikacijos. Ūgliai gali kauptis stiebo parenchimoje arba palikti įvairių žydėjimui ar sausringoms sąlygoms reikalingų medžiagų. Sandėliavimo ūgliai dažniausiai būna po žeme ir turi žvynuotus lapus bei atsinaujinančius pumpurus. Dėl to augalai tampa daugiamečiais, gali daugintis tokiomis sąlygomis, kai sunku sėti, ir užimti naujas teritorijas. Kartais vietoj lapų fotosintezės funkciją atlieka stiebai. Ūgliai gali virsti ūseliais ar dygliukais.
Skyrius Žalieji dumbliai – Chlorophyta.
Bendras rūšių skaičius – apie 15 tūkst.. Jos paplitusios visur, daugiausia gėlo vandens telkiniuose, kai kurios – jūrose ir labai mažai, esant periodinei drėgmei dirvožemyje, medžių kamienuose, tvorose, gėlių vazonuose ir kt.
Šio skyriaus atstovų pavyzdžiu galima atsekti dvi raidos kryptis: nuo vienaląsčių vienabranduolių formų iki sifoninių daugiabranduolių, aukščiausia šios linijos stadija yra caulerpa (Caulerpa gentis); nuo vienaląsčių formų iki kolonijinių iki daugialąsčių siūlinių ir toliau iki daugialąsčių su daugiau ar mažiau diferencijuotu talu, imituojantis išvaizda aukštesniųjų augalų organai, aukščiausia šios linijos stadija yra hara (Chara gentis).
Judančių formų judėjimo organai yra dvi, rečiau keturios vienodo ilgio ir formos žvyneliai. Ląstelės yra vienabranduolės, bet gali būti ir daugiabranduolės (Cladophora – Cladophoraceae šeima). Chloroplastai daugeliu atvejų su pirenoidais skiriasi savo forma, dydžiu ir skaičiumi ląstelėje. Pigmentai – chlorofilas, karotenoidai. Atsarginiai produktai – krakmolas ir aliejus. Dauginimasis yra vegetatyvinis, nelytinis ir seksualinis. Lytinis procesas žinomas beveik visoms rūšims ir yra labai įvairus: izogamija, heterogamija, oogamija, somatogamija (hologamija, konjugacija).
Žalieji dumbliai skirstomi į tris klases: lygias žiuželes, konjugatus ir anglį.
Isoflagellates klasė – Isocontophyceae.
Dauguma didelė klasė pagal rūšių skaičių. Talis yra vienaląsčiai, kolonijiniai, daugialąsčiai. Gyvenimo cikle yra daugiau ar mažiau ilga judrioji fazė.
Chlamydomonas (Chlamydomonas gentis). Šios genties rūšys dažniausiai gyvena sekliuose užterštose rezervuaruose ir balose ir dažnai sukelia vandens „žydėjimą“. Tai vienaląsčiai įvairių formų dumbliai: apvalūs, ovalūs, kiaušiniški. Siena yra pektino-celiuliozės. Priekiniame gale yra dvi citoplazminės žvyneliai. Chloroplastas yra puodelio formos, su įgaubtu paviršiumi, nukreiptu į priekinį ląstelės galą. Bazinėje chloroplasto dalyje yra gana didelis pirenoidas, apsuptas atsarginių krakmolo granulių, o viršutinėje dalyje yra stigma („akis“). Citoplazmoje, užpildančioje chloroplasto įdubą, yra branduolys, o žvynelinės apačioje – pulsuojanti vakuolė.
Palankiomis sąlygomis chlamidomonas dauginasi aseksualiai: protoplastas dalijasi mitotiškai į dvi, keturias ar aštuonias dalis, iš kurių net motininėje ląstelėje susidaro zoosporos, bendra struktūra identiškos suaugusiems, bet mažesnio dydžio ir be celiuliozės sienelės. Dėl motininės ląstelės sienelės lieknėjimo jos išsiskiria, užauga iki suaugusiųjų dydžio ir susiformuoja nauja ląstelės sienelė. Trūkstant vandens ir deguonies, chlamidomonas išmeta žvynelius ir išskiria gleives. Tuo pačiu metu protoplastas išlaiko gebėjimą dalytis. Susidarius palankioms sąlygoms, naujai susidariusios ląstelės suformuoja žiuželius, išsiskiria iš gleivių ir užauga iki normalaus dydžio. Seksualinis procesas dažnai būna izogamiškas, tačiau kai kuriose rūšyse buvo pastebėta heterogamija ir net oogamija. Susidariusi zigota užpildoma atsarginiais produktais ir sukuria storą sienelę. Tada ateina poilsio laikotarpis. Esant palankioms sąlygoms, zigotos turinys dalijasi mejozės būdu, todėl susidaro keturios haploidinės zoosporos.
Chlorella (Chlorella gentis). Šios genties rūšys plačiai paplitusios gėlo vandens telkiniuose, jūrose, dirvožemyje, ant medžių kamienų žievės. Kartais jie yra kerpių dalis. Talis yra vienaląstis. Ląstelė yra apvalios formos, savo sandara panaši į chlamidomonas, tačiau be žvynelių ir pulsuojančių vakuolių. Sporos neturi žvynelių. Jie vadinami aplanosporomis. Motininėje ląstelėje susidaro aštuonios sporos, kurios, augdamos, išleidžiamos ir pasyviai nešiojamos vandens srauto. nelytinis dauginimasis chlorella yra labai greita. Lytinio proceso nėra (kitų šaltinių duomenimis, kai kurios rūšys jį turi). Chlorelės ląstelės sukaupia daug atsarginių produktų, vitaminų, antibiotikų, todėl auginamos įvairioms reikmėms.
Ulothrix (Ulothrix gentis). Šios genties rūšys paplitusios upėse. Talis yra siūlinis, neišsišakojęs, susideda iš vienos eilės identiškų ląstelių, auga viršūnėje ir prie substrato yra pritvirtintas bespalve bazine ląstele. Chloroplastas yra žiedo arba pusžiedžio formos ir užima parietalinę padėtį. Šerdis yra viena. Nelytinio dauginimosi metu bet kurioje ląstelėje, išskyrus bazinę, susidaro keturviečiai zoosporos. Seksualinis procesas yra izogamiškas. Gametos yra mažos, dvisluoksnės, taip pat susidaro bet kurioje iš ląstelių. Susilieja tik skirtingų individų gametos (heterotalizmas). Zigota dalijasi mejozės būdu. Dėl to susidaro keturios haploidinės zoosporos, kurios sudygsta į suaugusius siūlus. Visas gyvenimo ciklas vyksta haploidinėje fazėje, tik zigota yra diploidinė.
Caulerpa (Caulerpa gentis). Šios genties rūšys yra jūros dumbliai, kurių sifoninis talas, šliaužiantis palei substratą, iki 50 cm ilgio, o kartais ir daugiau. Išoriškai jis primena šakniastiebį su papildomomis šaknimis ir dideliais lapais. Tai tarsi viena milžiniška ląstelė su vienu protoplastu, daugybe branduolių ir chloroplastų. Talio ertmė neturi pertvarų, bet yra kertama celiuliozės atraminėmis virvėmis. Tiesą sakant, nelytinio dauginimosi nėra, kartais vyksta vegetatyvinis dauginimasis talo dalimis. Seksualinis procesas yra izogamiškas. Visas gyvavimo ciklas vyksta diploidinėje fazėje. Mejozė atsiranda prieš formuojantis izogametams.
Klasės konjugatai – Conjugatophyceae.
Talis yra daugialąstis, siūlinis arba vienaląstis be žvynelių. Seksualinis procesas somatogamijos (konjugacijos) forma. Nėra zoosporų ar gametų.
Spirogyra (Spirogyra gentis). Daugybė šios genties rūšių gyvena gėlame vandenyje – upėse, tvenkiniuose, ežeruose ir durpynuose. Gijinis talis susideda iš vienos ląstelių eilės. Chloroplastai, 1-2 vienoje ląstelėje, yra citoplazmos sienelės sluoksnyje. Jie atrodo kaip spirališkai susuktos juostelės su pirenoidais, juostelių kraštai dažnai būna dantyti. Branduolys yra ląstelės centre ir yra panardintas į citoplazmą, kurios ploniausi siūlai tęsiasi iki jos sienelės sluoksnio. Kelios vakuolės. Spirogyra auga dalijantis ląstelėms. Vegetatyvinis dauginimasis vyksta talio fragmentais. Seksualinis procesas vyksta taip: lygiagrečiai išsidėstę du heterotaliniai individai; jų ląstelėse atsiranda sienų išsikišimų, kurie auga vienas kito link; sandūroje sienelės nusileidžia, susidaro konjugacijos kanalas, kuriuo protoplastas iš vieno individo, sąlyginai patino, ląstelės patenka į patelės ląstelę. Seksualinis procesas baigiasi susidarius didelei sferinei zigotai, kuri gamina storą sienelę ir atsarginius produktus aliejaus pavidalu. Po ramybės periodo zigota dalijasi mejozės būdu. Tokiu atveju susidaro keturios haploidinės ląstelės, trys iš jų miršta, o iš vienos išdygsta naujas individas. Taigi gyvavimo ciklas vyksta haploidinėje fazėje, tik zigota yra diploidinė.
Characeae klasė – Charophyceae.
Dideli dumbliai su sudėtingai išpjaustytu talu. Jie dažniausiai gyvena vandens telkiniuose su gėlo vandens(ežerai, šlaunies ežerai), kur formuoja tankius krūmynus. Zoosporomis nelytinio dauginimosi nėra. Vegetatyvinį atlieka specialūs „mazgeliai“, susidarę ant rizoidų arba talo dalių. Lytinio dauginimosi organai – oogonija ir anteridijos – yra daugialąsčiai. Characeae yra evoliuciniu požiūriu labiausiai pažengę žalieji dumbliai.
Hara (Chara gentis). Šios genties rūšyse talis siekia keliasdešimt centimetrų ilgio. Jis tarsi padalintas į „mazgus“ ir „tarpmazgius“, šakos nukrypsta nuo „mazgų“. Ašinę talio dalį sudaro vidutinė didelė ilga ląstelė, kurią supa mažesnės. Ilgos ląstelės išilgai talio kaitaliojasi su trumpesnėmis. Rizoidų pagalba talis pritvirtinamas prie rezervuaro dugno.
Vegetatyvinį dauginimąsi vykdo „mazgeliai“, susidarę ant rizoidų. Lytinio dauginimosi metu kai kurių šoninių vienaląsčių šakų sinusuose susidaro oogonijos ir anteridijos. Oogonia yra pailgos sferinės formos. Jo sienelė susideda iš spirališkai susuktų pailgų ląstelių, kurios viršuje baigiasi penkiomis trumpomis ląstelėmis (karūna). Viduje yra kiaušinis. Anteridijos yra mažesnės nei oogonijos ir turi sferinę formą. Subrendę jie būna oranžinės spalvos. Anteridiumo siena susideda iš aštuonių trikampių ląstelių - skruostų. Iš kiekvieno skydo į vidų išeina ilga ląstelė (rankena), kurios viršuje (galvoje) yra sferinė ląstelė, kuri sudaro spermatozoidinius siūlus. Pastarųjų ląstelėse susidaro spermatozoidai su dviem vienodais žvyneliais. Apvaisintas kiaušinėlis išauga į zigotą (oosporą), kuri pereina į ramybės periodą. Prieš daigumą atsiranda mejozė. Tada susidaro haploidinis trumpas nešakotas siūlas - priešaugis, iš kurio išauga naujas augalas. Gyvenimo ciklas vyksta haploidinėje fazėje, tik zigota yra diploidinė.
Paskaita Nr.9
Aukštesni sporiniai augalai.
aukštesni augalai.
Daugumoje aukštesnių augalų kūnas skirstomas į organus – šaknį, stiebą ir lapus, susidedančius iš gerai izoliuotų audinių. Aukštesniųjų augalų gyvenimo cikle aiškiai išreikšta sporofito (2n) ir gametofito (n) kaita. Lytinio dauginimosi organai yra daugialąsčiai. Patelė – archegonija – susideda iš išsiplėtusios apatinės dalies – pilvo, kuriame formuojasi kiaušinėlis, o viršutinės susiaurėjusios – kaklelio, kuris atsiveria kiaušiniui subrendus. Vyriškas lytinio dauginimosi organas – anteridiumas – atrodo kaip maišelis, kurio viduje susidaro daug spermatozoidų. Gimnosėkliuose sumažėjo anteridijos, o gaubtasėkliuose sumažėjo ir anteridijos, ir archegonijos. Iš zigotos aukštesniuose augaluose susidaro embrionas – sporofito gemalas.
Bryophyta skyrius – Bryophyta.
Bendras rūšių skaičius – apie 35 tūkst.
Struktūra. Briofitų, kaip ir kitų aukštesnių augalų, gyvavimo cikle kaitaliojasi dvi fazės: sporofitas ir gametofitas. Tačiau dominuoja (vyrauja) gametofitas, o visuose kituose aukštesniuose augaluose dominuoja sporofitas. Štai kodėl aukštesniųjų augalų evoliucijoje bryofitai laikomi nepriklausoma šonine šaka.
Bryofitai savo organizacija ir ekologija vis dar artimi dumbliams. Kaip ir dumbliai, jie neturi indų ir šaknų. Kai kurie primityvūs atstovai turi augalinį kūną šliaužiančio talo pavidalu su viršūniniu (dichotominiu) išsišakojimu, panašų į dumblių talą. Tręšimas yra susijęs su vandeniu. Tarp briofitų, taip pat tarp dumblių, lignified formų nėra.
Sklaidymas. Bryofitai paplitę visuose pasaulio žemynuose, tačiau netolygiai. Atogrąžų šalyse – daugiausia kalnuose. Nedidelis skaičius rūšių auga sausringomis sąlygomis, pavyzdžiui, stepėse. Kai kurios rūšys gyvena epifitiškai ant medžių ar vandens žievės. Pagrindinė rūšių įvairovė telkiasi drėgnose šiaurinio pusrutulio vietose, vidutinio ir šalto klimato zonose. Jie vaidina svarbų vaidmenį formuojant augalijos dangą, ypač tundroje, pelkėse ir miškuose.
klasifikacija. Bryofitai skirstomi į tris klases: anthocerotes, kepenėles, lapines samanas. Aukščiausia vertė turi paskutines dvi klases.
Klasė Liverworts - Hepaticopsida.
Bendras rūšių skaičius apie 10 tūkst.. Jos paplitusios visur. Primityvi kepenėlių kūno sandara byloja apie jų senumą.
Paprastoji marčantija (Marchantia polymorpha) yra tipiškas klasės atstovas. Gametofitas sluoksninio talo pavidalo, 10-12 cm ilgio, viršūniškai išsišakojęs. Iš abiejų pusių jis padengtas epidermiu. Viršutiniame epidermyje yra ventiliacijos angos – stomos. Juos supa specialūs narvai, išdėstyti keturiomis eilėmis. Po stoma yra oro kameros. Apatiniame epidermyje susidaro ataugos – vienaląsčiai šakniastiebiai ir rausvos arba žalsvos spalvos žvyneliai, kurie kartais painiojami su sumažėjusiais lapais. Po viršutiniu epidermiu yra asimiliacinis audinys, susidedantis iš vertikalių parenchiminių ląstelių stulpelių su chloroplastais. Žemiau yra plonasienių chlorofilo neturinčių parenchiminių ląstelių sluoksnis. Vadinasi, marchantia tallus turi dorsiventralinę struktūrą.
Viršutinėje talo pusėje susidaro specialios šakos - padėkliukai, o ant jų - lytinio dauginimosi organai. Marchantia yra dvinamis augalas. Kai kuriuose egzemplioriuose stovai yra devynių spindulių žvaigždės, sėdinčios ant kojos, formos, tarp kurios spindulių apatinėje pusėje yra archegonijos. Kitose atramos yra aštuonkampio skydo, sėdinčio ant kojos, pavidalo, kurio viršutinėje pusėje yra anteridijos, panardintos į anteridines ertmes. Archegonijos pilve susidaro kiaušialąstė. Po jo susiliejimo su sperma iš zigotos susidaro sporogonas. Tai dėžutė ant trumpo kotelio, kurią prie gametofito pritvirtina haustorija. Dėl mejozės dėžutės viduje iš sporogeninių ląstelių susidaro haploidinės sporos, taip pat elateriai – negyvos pailgos ląstelės su spirale sustorėjusia sienele, kurios padeda supurenti sporų masę, taip pat jas išmesti iš dėžutės. . Esant palankioms sąlygoms, iš sporos išsivysto priešaugis arba protonema. Tai mažas siūlas. Marchantia thallus auga iš savo viršūninės ląstelės.
Vegetatyvinį dauginimąsi atlieka lęšio formos, žalios spalvos perų kūnai. Jie susidaro viršutinėje talijos pusėje specialiuose krepšeliuose dėl jų dugną išklojusių ląstelių dalijimosi.
Marchantia rūšys yra plačiai paplitusios. Dažniausiai juos galima rasti drėgnose vietose: ežerų ir upių pakrantėse, daubose ir žolinėje padangėje po miško laja.
Klasė Lapinės samanos – Bryopsida.
Bendras rūšių skaičius – apie 25 tūkst.. Daugelis rūšių yra paplitusios Šiaurės pusrutulio cirkumpoliarinėse šalyse. Didelėse tundros, pelkių ir miškų teritorijose jie dominuoja augalijos dangoje, o tai daro didelę įtaką drėgmės tiekimui į žemę.
Gametofitas yra stačios, panašios į stiebą ašis – kaulidiumas, padengtas lapo formos ataugomis – filidijomis. Tradiciškai jie gali būti vadinami stiebu ir lapais. Apatinėje stiebo dalyje susidaro daugialąsčiai rizoidai (ne visi). Šoninis išsišakojimas. Ašių augimas atsiranda dėl piramidinės viršūninės ląstelės dalijimosi. Jis gali būti monopodinis arba simpodinis. Pagal tai lytinio dauginimosi organai ir sporogonas yra gametofito viršuje arba ant šoninių šakų.
Klasė skirstoma į tris poklasius: Andreevy samanos, Sphagnum samanos, Brie (žaliosios) samanos. Paskutiniai du poklasiai turi didžiausią reikšmę.
Poklasis Sphagnum samanos – Sphaqnidae.
Sfagninės samanos turi gana vienodą struktūrą, todėl jas sunku atpažinti. Jų gametofitas yra stipriai šakojantis augalas, ypač viršutinėje dalyje. Šakos tankiai padengtos lapais. Sfagninės samanos gyvena labai drėgnoje aplinkoje. Šiuo atžvilgiu jie neturi rizoidų, o drėgmė patenka tiesiai į stiebą, kuris laikui bėgant miršta prie pagrindo. Stiebo struktūra paprasta. Jo centre yra plonasienių parenchiminių ląstelių šerdis, kuri atlieka laidumo ir saugojimo funkcijas. Jį gaubia žievė, susidedanti iš dviejų sluoksnių: sklerodermos, atliekančios mechaninę funkciją, ir hialodermos, atliekančios vandens kaupimo funkciją. Hialodermos ląstelės yra didelės, negyvos, jų sienelėse yra apvalios skylės, per kurias susisiekia gretimų ląstelių ertmės tarpusavyje, taip pat su išorinė aplinka. Kartais šios ląstelės turi spiralinį sustorėjimą. Lapas susideda iš vienos eilės ląstelių, kurios labai skiriasi tiek struktūra, tiek savo funkcijomis. Vienos gyvos, turinčios chlorofilą, kitos – negyvos, santykinai didesnės, spirale sustorėjusiomis sienelėmis, pradurtos skylutėmis, savo sandara panašios į vandenį kaupiančias hialodermos ląsteles, vadinamos hialininėmis. Hialino ląstelės sugeba sukaupti ir ilgą laiką išlaikyti didžiulį vandens kiekį, 30–40 kartų didesnį už paties augalo masę.
Gametofitai yra vienanamiai ir dvinamiai. Anteridijos susidaro lapų pažastyse ant stiebo atšakų. Aplink juos lapai nudažyti rausva spalva. Archegoniumas ant trumpų šakų. Dėl spermatozoidų susiliejimo su kiaušialąste susidaro zigota, kuri reiškia diploidinės fazės pradžią - sporogoną. Sporogonas susideda iš stiebo ir kapsulės. Kotelis labai sutrumpėjęs, svogūninis, tačiau sporoms subrendus, gametofito stiebo viršūnė stipriai išauga ir iškelia dėžutę (netikras stiebas). Dėžutės centre dedamas suapvalintas stulpelis, virš kurio dedamas sporangijų rinkinys su sporogeniniu audiniu. Dėžutės sienelė tvirta, daugiasluoksnė. Išoriniame chlorofilo sluoksnyje yra didelis skaičius neišsivysčiusios stomos. Dėžutė turi dangtelį, kuris brendimo metu atšoka ir sporos išsisklaido. Elater nėra. Pirmiausia iš sporų susidaro žalia lamelė protonema, o paskui iš ant jos esančių pumpurų – suaugęs gametofitas, kuris dominuoja gyvavimo cikle.
Sphagnum struktūra yra primityvi: plokščia protonema, kraujagyslių pluošto ir rizoidų nebuvimas, silpna kapsulės diferenciacija.
Sfagnų vertė gamtoje yra labai didelė. Sukaupdamos didžiulį vandens kiekį ir išaugdamos į tankias velėnas, jos sukelia didžiulių erdvių užpelkėjimą, pasiekiančią tundros zoną. Jiems nusausinti atliekami agromelioracijos darbai. Kita vertus, senos pelkės turi didelę ekonominę reikšmę durpių telkinių plėtrai. Durpių sluoksnio augimas palankiausiomis sąlygomis vyksta lėtai – apie 10 metų susidaro 1 cm storio sluoksnis.
Poklasis Brie (žalios) samanos – Bryidae.
Rūšių skaičius – 24,6 tūkst.. Jos paplitusios plačiau nei sfagninės samanos. Jie gyvena įvairiomis aplinkos sąlygomis – nuo tundros ir miško-tundros iki stepių ir dykumų. Tipiškiausios dygliakinių samanų buveinės, kuriose jos dominuoja arba sudaro ištisinę dangą, yra tundra, pelkės ir kai kurių rūšių miškai. Kiekviena buveinė turi savo rūšis. Brie samanos, palyginti su sfagninėmis samanomis, išsiskiria didele struktūrų įvairove. Vienų rūšių lytinio dauginimosi organai yra išdėstyti ant pagrindinės ašies, kitų - ant šoninių. Kai kuriose rūšyse šakojimasis nėra išreikštas.
Paprastieji gegutiniai linai (Polytrichum commune) yra vienas iš dažniausių žiobrių samanų atstovų. Auga miškuose, laukymėse, pelkių pakraščiuose.
Gametofito stiebas stačias, nešakotas, 15 cm ir daugiau aukščio, tankiai apaugęs lapais. Požeminė jo dalis dirvoje tęsiasi beveik horizontaliai, ant jos susidaro rizoidai. Stiebo centre yra koncentrinis kraujagyslių pluoštas, susidedantis iš pailgų ląstelių, panašių į tracheidus ir sieto vamzdelius. Jį supa parenchima, kuri atlieka ir laidumo funkciją. Iš išorės parenchima ribojasi su sklerodermija (žieve). Jo išorinis sluoksnis, susidedantis iš bespalvių ląstelių, vadinamas hialodermija.
Lapai išsidėstę spirale. Jie susideda iš linijinės plokštės su smailia dantyta viršūne ir membraniniu apvalkalu. Asimiliacinės plokštės yra viršutinėje lapo pusėje. Išplečiama vena su mechaniniais ir laidžiais elementais.
Gametofitas yra dvinamis. Butelio formos archegonijos yra patelės gametofito viršuje, maišo formos anteridijos – patino viršuje. Tarp archegonijos ir anteridijų yra sterilūs siūlai – parafizės. Po apvaisinimo iš zigotos susidaro sporogonas, susidedantis iš ilgo stiebo ir dėžutės. Kapsulė yra stačia arba daugiau ar mažiau įstriža, prizminė, keturių ar penkių pusių, padengta rūdžių veltinio gaubteliu, suformuotu iš archegonijos sienelių. Dėžutė susideda iš urnos ir dangčio. Apatinė dalis urna susiaurėjusi ties kaklu. Epidermyje ant urnos ir kaklo ribos yra stomos. Urnos centre yra kolona, kuri ties dangteliu išsiplečia ir suformuoja epifragmą – plonasienę pertvarą, kuri uždaro urną. Aplink koloną yra cilindrinio maišelio pavidalo sporangija, pritvirtinta prie sienos ir kolonos specialiais siūliniais dariniais. Urna turi specialų prietaisą sporoms išsklaidyti – peristomą, tai yra gvazdikėlių eilė bukomis viršūnėmis, išsidėsčiusi palei urnos kraštą. Tarp dantų, galinčių atlikti higroskopinius judesius, ir epifragmos yra angos, pro kurias esant sausam orui išsilieja sporos. Iš sporos išauga protonema žalio išsišakojusio siūlo pavidalu. Ant jo susidaro inkstai, iš kurių laikui bėgant išsivysto suaugę gametofitai.
Skyriai Rhynioid - Rhyniophyta ir Psilotoid - Ps1lotophyta.
Rhyniform skyrius apima 2–3 tik iškastinių augalų gentis. Gyvenimo cikle dominuoja sporofitas. Jo vegetatyvinis kūnas susideda iš šakotų kūnų sistemos. Bendra struktūra anteninėje kūno dalyje yra labai savotiška. Tai dar ne ūglis, nes ant telomo ašių nėra lapų. Pagrindinė ašis yra gerai apibrėžta. Išsišakojimas yra viršūninis (dichotominis). Ašies centre ksilemas yra izoliuotas, apsuptas floemo. Ksilemas gali būti kompaktiškai išdėstytas cilindro arba sijų pavidalu. Jį sudaro tracheidai. Periferinė (žievės) kūno dalis atlieka fotosintezės funkciją. Epidermyje yra stomatalinis aparatas. Požeminėje dalyje stomų nėra. Tikrų šaknų nėra, jas pakeičia rizoidai. Sporangijos išsidėsčiusios telomos viršūnėse, sporangijos sienelė daugiasluoksnė. Rhiniformes gametofitų nerasta. Atstovas yra Rhynia gentis, kuriai priklauso dvi rūšys. Tai apie 20 cm aukščio, 3 mm skersmens žoliniai augalai. Požeminė dalis susideda iš horizontalaus korpuso, iš kurio statmenai tęsiasi oro ašys.
Šiuolaikinėje floroje Psilotoidų departamentui priklauso dvi gentys: psilo (Psilotum) ir tmesipter (Tmesipteris). Bendras rūšių skaičius yra 4 - 6. Abi gentys yra plačiai paplitusios abiejų pusrutulių atogrąžų ir subtropikų zonose.
Psilotoidų sporofitas yra epifitinis, retai sausumos žolinis augalas. Kūno ilgis 5 - 40 (iki 100) cm.Šakojimai dažnai būna viršūniniai. Žievė gerai išsivysčiusi, atlieka fotosintezės funkciją. Stomatalinis aparatas yra primityvus. Lapai smulkūs, 1-5 mm ilgio, subuluoti, plokšti, be stuburėlių. prietaisai ir venos. Jie gali būti laikomi telomos ataugomis. Požeminę dalį vaizduoja šakniastiebis su rizoidais. Nėra šaknų. Sporangijos auga kartu po 2-3 (sinangijos), atviros išilginiu plyšiu. Tokio pat dydžio sporos. Psilotoidų sporofito struktūra rodo artumą rinoidams.
Gametofitas dvilytis, be chlorofilo, radialiai simetriškas, viršūniškai išsišakojęs. Jo ilgis apie 20 mm, skersmuo 2 mm Saprofitiškai minta grybų pagalba, su kuriais patenka į simbiozę. Paviršius padengtas rizoidais. Gyvena daugiausia po žeme. Tręšimas yra susijęs su vandeniu.