Пресный водоём и его обитатели. Мхи — древние и важные
на тему: "Низшие и высшие растения: водоросли, мохообразные и папортникообразные"
Низшие и высшие растения
Растения делят на две группы, или два подцарства: низшие растения и высшие. Низшими называют такие растения, вегетативное тело которых не расчленено на органы, а высшими - растения со специализированными вегетативными органами, состоящими из разных тканей.
Систематика растений
Разобраться в биоразнообразии позволяет наука систематика. Существует систематика растений, животных, грибов, бактерий.
Собственно, в обычной жизни человек тоже систематизирует. Так, чашка, стакан, сахарница объединяются в категорию "чайная посуда", а тарелка, вилка, столовая ложка - в категорию "столовая посуда". Обе категории объединяются в более широкую категорию "посуда". Вместе с мебелью посуда, например, может быть объединена в еще более широкую категорию "домашняя утварь" и т.д.
Чем меньше ранг систематической категории, тем большее их число существует. Отделов растений известно всего 15-16, а видов - около 350 000. В одно семейство может входить 1000 родов. Известны роды в 2000-3000 видов. Но есть роды всего с одним видом и семейства с одним родом.
Вид - основная единица систематики. Называя какое-либо растение, мы подразумеваем, как правило, вид. Растения одного вида могут давать потомство, разные виды, как правило, не могут. Поэтому виды, обитающие в природе совместно, не скрещиваются и хорошо различаются. Каждый вид обязательно относится одновременно и ко всем остальным систематическим категориям. Например: осина относится к роду тополь, семейству ивовых, порядку ивоцветных, классу двудольных, к отделу цветковых растений.
Название вида состоит обычно из двух слов, причем первое слово обозначает род, а второе - собственно вид (например, береза бородавчатая, лютик едкий, смородина черная; исключение - осина, помидор, картофель).
В научных книгах названия обязательно пишутся и на латинском языке (что понятно ученым всего мира).
У культурных растений, в пределах одного вида, часто существует множество сортов. У домашней яблони, например, их несколько тысяч. Сорт - результат деятельности человека. Сорта в отличие от видов могут скрещиваться друг с другом.
Водоросли: их экология и значение
Общие особенности водорослей. Водоросли от других растительных организмов отличаются следующими признаками:
Водоросли, или низшие растения
Живут главным образом в воде.
Не имеют вегетативных органов.
Органы размножения одноклеточные.
Окраска разнообразна.
Высшие растения
Живут на суше (обитание в воде вторично).
Имеют вегетативные органы.
Органы размножения многоклеточные.
Листья всегда зеленые.
Водоросли могут быть одноклеточные и многоклеточные, иногда крупных размеров, до нескольких десятков метров. Они фотосинтезируют и имеют хлорофилл, но нередко и другие пигменты. Окраска водорослей может быть зеленая, желтоватая, бурая, красная. Клетка водорослей имеет те же части, что и у высших растений. Их фотосинтезирующие пластиды, от которых зависит окраска, называют хроматофора-ми. Размножаются водоросли вегетативным, бесполым и половым путем.
Зеленые водоросли, обитающие в пресных водоемах. Большинство зеленых водорослей живет в пресных водоемах, в морях их сравнительно немного. Существуют одноклеточные и многоклеточные водоросли.
Хламид о, монада и хлорелла. Мелкие одноклеточные водоросли. Их можно рассмотреть только при помощи микроскопа. Они имеют оболочку, ядро, цитоплазму, чашевидный хроматофор. Но отличаются существенными признаками.
Хламидомонада
Активно передвигается посредством жгутиков.
Имеется красный светочувствительный глазок.
Имеются пульсирующие вакуоли.
Бесполое размножение посредством подвижных спор со жгутиками - зооспор.
Половой процесс протекает при помощи двужгутиковых гамет.
Обитает в мелких пресных водоемах.
Хлорелла
Неподвижна.
Красный светочувствительный глазок отсутствует.
Пульсирующие вакуоли отсутствуют.
Бесполое размножение посредством неподвижных спор.
Половой процесс отсутствует.
Обитает в основном на сырой почве и стволах деревьев.
Спирогира. Самая обыкновенная нитчатая зеленая водоросль. Образует основную массу скользкой зеленой тины в прудах. Клетка имеет следующие части: оболочку, покрытую слизью; крупное ядро с ядрышком (хорошо заметное под микроскопом); цитоплазму; хроматофор в виде одной или нескольких лент, расположенных по спирали; вакуоль, занимающую большую часть клетки.
Вегетативное размножение у спирогиры происходит простым разрывом нитей. Споры бесполого размножения отсутствуют. Половой процесс - конъюгация, т.е. слияние обычных вегетативных клеток, а не особых гамет. Этапы конъюгации:
1 - две нити располагаются параллельно;
2 - противолежащие клетки дают выросты навстречу друг другу, так что образуется подобие лестницы;
3 - оболочки на концах отростков растворяются;
4 - содержимое одной из клеток перетекает в клетку напротив и сливается с ее содержимым, в результате чего образуется зигота.
Кладофора представляет собой не слизистые на ощупь ветвящиеся нити, прикрепленные к субстрату. Хроматофор - в виде сеточки. Каждая клетка имеет много ядер (это встречается иногда у водорослей, но никогда - у высших растений). Бесполое и половое размножение происходит примерно как у хламидомонады.
Таким образом, признаки четырех родов зеленых водорослей таковы:
Морские водоросли. Одни морские водоросли микроскопические, часто одноклеточные. Много их обитает в поверхностных слоях воды и составляют часть планктона. Другие обитают на дне, главным образом на камнях и подводных скалах, на сравнительно небольших глубинах (150 - 200 м), т.е. в основном в прибрежной полосе.
Водорослям необходим свет, поэтому на очень больших глубинах они существовать не могут. Их мало и там, где вода бедна питательными веществами. Основную массу донных водорослей составляют бурые и красные водоросли. Форма этих водорослей очень разнообразна: в виде кустиков, пластиночек, шнуров. Бурые водоросли окрашены в бурый, коричневый или почти черный цвета; красные - в розовый, ярко - или темно-красный. Бурые водоросли достигают самых крупных среди морских водорослей размеров. К ним относится, в частности, ламинария, или морская капуста.
Тело ламинарии (слоевище) напоминает длинный довольно узкий лист на черешке. Ко дну она прикрепляется выростами - ризоидами. Как и у других водорослей, ризоиды служат лишь для прикрепления: вода всасывается всей поверхностью. Ламинария достигает нескольких метров в длину. Внутреннее строение ее довольно сложное. У нее имеются даже ситовидные клетки, напоминающие ситовидные трубки высших растений. Но сосуды отсутствуют, поскольку у водорослей в них нет и надобности. У ламинарии образуются зооспоры, из которых вырастают микроскопические заростки с половыми органами. Так что цикл развития ламинарии несколько напоминает папоротники.
В прибрежной зоне наших северных морей обитает фукус, также бурая водоросль. Слоевище фукуса сильно рассечено на ремневидные доли. Оно гораздо мельче, чем у ламинарии (до 50 см длиной). Половые органы образуются в специальных вместилищах. Спор бесполого размножения нет. Значение морских водорослей в основном заключается в следующем:
планктонные водоросли играют большую роль в питании морских животных;
заросли донных водорослей дают приют рыбам и другим животным;
ламинарию и другие водоросли использует в пищу человек;
из бурых и красных водорослей получают йод и агар-агар;
хлорелла используется в космонавтике для восстановления нормального состава воздуха.
Мохообразные
Общие признаки. Мохообразные - растения, часто очень маленькие, сравнительно простого строения. В отличие от водорослей у них, как правило, имеются листья и стебли. Корни всегда отсутствуют; есть только ризоиды. Половые органы и спорангии многоклеточные. Цикл развития совершенно особенный - коробочки со спорангиями развиваются из зиготы непосредственно на растении.
Строение мохообразных. Зеленые, или бриевые, мхи. Последний эпитет удачнее, поскольку все мохообразные - зеленые растения.
Среди бриевых мхов один из самых крупных представителей - кукушкин лен. Его стебли достигают длины 20 см (для мхов это очень много). Стебель неветвящийся, густо покрыт узкими листьями, несколько напоминая настоящий лен (отсюда и название). Вместо корней - просто устроенные ризоиды, отходящие от нижней части стебля. Они служат как для прикрепления, так и для всасывания воды (в отличие от водорослей). По сравнению с водорослями бриевые мхи отличаются и сложным внутренним строением. Например, у кукушкина льна имеется подобие эпидермиса и проводящей ткани.
Кукушкин лен - двудомное растение: мужские и женские половые органы находятся на разных экземплярах, близ верхушки. Мужские половые органы - антеридии представляют собой мешочки, в них образуются сперматозоиды. Женские половые органы - архегоний похожи на колбочки с длинными шейками. Их стенка состоит из одного слоя клеток; в расширенной части колбочки находится яйцеклетка. Для оплодотворения необходимы дождь или роса. Тогда сперматозоиды могут попасть на архегоний и проникнуть через шейку к яйцеклетке. Из зиготы образуется коробочка на длинной ножке. Коробочка имеет крышечку и покрыта сверху колпачком. Внутри находится спорангий в виде муфты. В спорангии образуются споры, которые по созревании выпадают из коробочки. Для этого должна отпасть крышечка и разрушиться стенка спорангия. Ясно, что чем длиннее ножка, тем дальше могут рассыпаться споры. Спора прорастает, образуя тонкую зеленую нить. На нити появляются почки, из которых вырастают побеги мха.
Бриевые мхи очень распространены в природе. Их можно найти на болотах, лугах, пустынях. Особенно много их в тенистых лесах. Не все они похожи на кукушкин лен. У многих стебли сильно ветвятся, часто они стелющиеся. Немало мхов, у которых стебли не превышают 2-3 см. Различной формы могут быть и коробочки. Но жизненный цикл у всех одинаковый.
Торфяные, или сфагновые, мхи. Торфяные мхи растут на торфяных болотах, вместе с клюквой, голубикой и багульником. Лишь очень немногие растения уживаются с торфяными мхами. Они всегда возникают в массе, образуя сплошной ковер. Стебель сфагновых мхов ветвится, образуя веточки трех типов: одни отходят в стороны, другие свисают, прилегая к стеблю, третьи образуют подобие головки на верхушке. Листья очень мелкие (едва видны невооруженным глазом) и состоят из одного слоя клеток. Клетки встречаются двух типов: крупные водоносные, прозрачные, со спиральными утолщениями стенок и узкие хлорофиллоносные, зеленые. Каждая водоносная клетка окружена несколькими хлорофилл оносными клетками. Водоносные клетки могут очень быстро впитывать огромное количество воды (в 25 раз больше сухого веса) и столь же быстро ее терять. Благодаря такой особенности у сфагнума нет не только корней, но и ризоидов (они ему не нужны). Размножаются сфагновые мхи так же, как и бриевые.
Растения сфагнума нарастают сверху и отмирают снизу. Отмирающие нижние части вместе с другими растениями превращаются в торф. Последний образуется при неполном разложении частей растений (не хватает кислорода). Торф - ценное топливо. Однако во многих случаях осушение болот нежелательно. Во-первых, могут возникнуть изменения климата; во-вторых, на сфагновых болотах часто встречаются редкие растения. Целый ряд сфагновых болот получил сейчас статус памятников природы.
Папоротникообразные
Общие признаки. Папоротникообразные имеют корни и побеги (стебли с листьями). Размножаются они спорами. Половые органы образуются на особых мелких растеньицах - заростках.
Строение папоротников. Папоротники широко распространены. У них крупные, сильно рассеченные листья, отходящие от корневища. На корневище образуются и придаточные корни. Черешки покрыты буроватыми чешуйками. Верхушка молодых листьев свернута в улитку. В процессе роста улитка раскручивается, и лист растет верхушкой, как побег. За эту особенность листья папоротника называют иногда плосковетками.
Размножение папоротников. На нижней стороне листа (но не каждого) образуются спорангии, расположенные кучками и часто прикрытые покрывальцами или краем пластинки листа. Отдельный спорангий простым глазом увидеть трудно. Его строение идеально приспособлено для разбрасывания спор. По форме он похож на двояковыпуклую линзу. Стенки спорангия состоят из одного слоя клеток. Все они тонкостенные, за исключением клеток, расположенных по гребню (кольцо). У этих клеток утолщены внутренняя и боковые стенки. Важно, что кольцо занимает не весь гребень, а 2/3 его, стало быть, остается тонкостенная часть гребня. При созревании спор стенка спорангия разрывается, а кольцо, подобно пружине, разбрасывает споры. Из споры вырастает крошечное растеньице, в виде сердцевидной, прижатой к земле пластиночки. Это - заросток. У него есть ризоиды; на нижней стороне образуются антеридии и архегонии. Оплодотворение происходит как и у мохообразных. Из зиготы развивается зародыш, а потом и молодое растение папоротника.
Разнообразие папоротников. Папоротники - преимущественно лесные растения. Особенно много их во влажных лесах тропиков. Большинство их имеет сильно рассеченные листья, часто очень крупных размеров. Но есть немало папоротников и с цельными листьями. Некоторые представляют собой лианы с вьющимися стеблями или листьями, есть напоминающие деревья, со стволами 10 м и более высотой. Особенно много среди папоротников эпифитов, поселяющихся на стволах и ветвях деревьев. В умеренных широтах папоротников немного. Обычны у нас мужской папоротник, женский папоротник (названия восходят к глубокой древности, когда еще было неизвестно, как размножаются папоротники), орляк, страусопер и некоторые другие.
Хвощи и плауны. Это также многолетние травянистые споровые растения.
Их особенности в сравнении с папоротниками следующие:
Былой расцвет папоротникообразных. В группе папоротникообразных насчитывается 13 000 видов. Примерно 300 млн лет назад на Земле еще не было цветковых растений. Уже появились голосеменные, но особенно большую роль играли папоротникообразные. Многие из них были настоящие деревья, с камбием, достигавшие высоты 40 м, их стволы иногда были не меньше 1 м в диаметре. Некоторые напоминали хвощи, увеличенные до гигантских размеров, другие - плауны. Травы также были представлены исключительно папоротникообразными и мохообразными. Климат был теплый и влажный, а освещение - менее интенсивное, чем сейчас. Леса часто были заболочены, погибая, деревья падали в воду, заносились илом. Постепенно стволы спрессовывались и без доступа кислорода превращались в каменный уголь - прекрасное топливо.
Растение - целостный организм
Органы растения - и вегетативные, и генеративные - находятся в сложной взаимосвязи, обеспечивая жизнь единого организма. Корни поглощают из почвы воду и минеральные соли, которые нужны для нормального существования всех живых клеток. В корнях образуются органические вещества: аминокислоты, витамины, гормоны, ферменты и другие соединения, без которых невозможна жизнь организма. Некоторые из них идут на образование в листьях хлорофилла. Без хлорофилла не происходит процесс фотосинтеза. Для фотосинтеза необходима вода, которая также поступает к зеленым клеткам листа из корней.
Большое количество воды испаряется надземными органами, и тем самым растение предохраняет себя от перегрева. Воду к побегам подают корни.
В свою очередь, в клетках корней синтез разнообразных жизненно важных соединений возможен при поступлении в них органических веществ из листьев. Только в клетках с хлоропластами образуются органические вещества из неорганических - воды и углекислого газа. Продукты фотосинтеза необходимы корням для их роста и ветвления.
Таким образом, только при тесной взаимосвязи между надземными и подземными вегетативными органами возможна жизнь организма.
Цветение, созревание плодов и семян также невозможно без обеспечения генеративных органов всеми необходимыми им веществами. Эти вещества им поставляют вегетативные органы.
В свою очередь, генеративные органы оказывают влияние на жизнедеятельность вегетативных органов. Так, работа корней зависит не только от органов воздушного питания, листьев, но и генеративных органов. В опытах было показано, что удаление завязей из ряда цветков пшеницы или затенение колосьев приводило к заметному уменьшению поступления азота из корней в надземную часть растения.
Приведенные примеры свидетельствуют о том, что растительный организм - это единая и целостная система. В этой системе функции поделены между отдельными органами, но деятельность их тесно взаимосвязана.
На протяжении суток или в течение года, изменение температуры при перемещении от экватора к полюсам и т.д. Для животных существование можно считать нормальным, если изменения экологического фактора находятся в пределах его максимального и минимального значений, то есть границ толерантности.
Между значениями, ограничивающими возможность существования животного, находится наиболее для него благоприятная, но относительно узкая зона оптимума. Между зоной оптимума и максимумом или минимумом находится зона пессимума, в которой на организм влияют неблагоприятные условия.
Зоны оптимума и пессимума устанавливаются по числу особей, приходящихся на единицу площади, то есть по плотности популяций вида.
Определение 1
Экологическая валентность вида – это способность данного вида заселять разную среду, которая характеризуется различными изменениями экологических факторов.
Стенобионтный вид – это вид с малой экологической валентностью, приспособленный переносить только ограниченные колебания факторов среды. Сухопутные представители: обитающая во влажных тропических лесах человекообразная шимпанзе, обитатель хвойного леса умеренной зоны – глухарь. Стенобионты водной среды: рифообразующие кораллы, для нормального роста и развития которых необходимы строго определенные температура, степень освещенности и соленость морской воды. Эти кораллы живут в тропической зоне Мирового океана.
Эврибионтный вид – это вид, способный заселять местообитания с меняющимися условиями, или различные местообитания (лисица – встречается в горах и на равнине, в степи и в лесу, в границах нескольких ландшафтно – климатических зон; киты могут существовать в различных широтах).
При указании относительной степени толерантности применяют соответствующие термины с приставкой «стено» - узкий или «эври» - широкий:
- относительные температуры – стенотермные или эвритермные;
- пищевой рацион – стенофаги или эврифаги;
- соленость воды – стеногалинны или эвригалинны;
- по отношению к месту обитания – стеноэки и эвриэки и т.д.
Условия распространения и существования животных в морской среде
Моря и океаны включают наиболее богатую фауну, на долю которой приходится 64% всех видов животных. Многие классы животных способны обитать только в море: плеченогие, коралловые полипы, головоногие и боко-нервные моллюски, оболочники, бесчерепные, губки, немертины, многощетинковые кольчецы и др.
На морские организмы особое влияние оказывают:
- давление;
- плотность;
- глубина проникновения солнечной радиации;
- растворение солей и содержание газов;
- распределение тепла;
- течения.
В морской среде выделяют две группы биотопов:
- Пелагиаль – толща морской воды, в которой жизнь не связана с твердым субстратом (берег, дно). Представители пелагиали активно или пассивно плавают (нектон, планктон, китообразные, рыбы, моллюски (кальмары), крупные головоногие).
- Бенталь – вся поверхность дна и берег. Для бентали характерно вертикальное расчленение, выделяют литоральную и абиссальную зоны. Резкой границы между этими зонами нет, их делит батиальная зона.
Литоральная зона включает поверхность континентального шельфа от зоны заплеска до начала материкового ската. Зона характеризуется наличием весьма разнообразных биотопов и обилием пищи.
Абиссальная зона состоит из морского дна и прилегающего слоя воды от края материкового ската. Наиболее однообразная среда обитания на планете. Пищевые ресурсы скудные, представлены в основном детритом. Главные обитатели на большой глубине – иглокожие. На дне обитают фильтраторы: черви, актинии, моллюски, морские пауки-пантоподы, крабы и др. животные. Имеется много реликтов – рыбы-химеры, стебельчатые морские лилии и некоторые другие представители.
Условия распространения и существования животных в пресных водах
Пресные водоемы разобщены и прямых связей между населяющих их животных, нет. В пресных водоемах нельзя встретить головоногих моллюсков, иглокожих, многощетинковых кольчецов, но могут попадаться представители кишечнополостных и губок. Только с пресными водоемами связаны земноводные животные.
Пресная фауна происходит от морской, поэтому на распространение организмов оказывают влияние общие факторы среды:
- Соленость водоемов. Некоторые виды животных – эвригалинные организмы - приспособились к колебаниям солености воды. Стеногалинные организмы к степени солености строго избирательны.
- Жесткость воды. Так, мшанки, губки, некоторые виды раков могут обитать только в мягкой воде.
- Содержание гумуса и растворенного кислорода. В водоемах, богатых гумусом, мало живых организмов. Разложение органических веществ ведет к заморам рыбы, что особенно опасно для стеноксибионтов.
- Температура. Большинство представителей внутренних вод относят к эвритермным организмам.
- Течения. Представители пресноводных водоемов чувствительны к скорости движения воды, их делят на реофилов – обитателей текучих вод и лимнофилов – обитатели стоячей воды.
Условия распространения и существования наземных животных
На суше происходят широкие колебания всех факторов среды обитания.
Главнейшие факторы, обуславливающие распространение сухопутных животных:
- Влажность воздуха и почвы. Различают гигрофилы – влаголюбивые животные и ксерофилы – сухолюбивые, есть организмы, существование которых не зависит от влажности.
- Движение воздуха. Наличие сильного ветра (острова, морские побережья) отпугивает летающих насекомых, а за ними исчезают и насекомоядные животные.
- Температура. Каждый вид имеет свой собственный диапазон температур – температурный оптимум вида, обеспечивающий благоприятное существование. Эвритемные виды в основном заселяют умеренные климатические пояса, стенотермные виды подразделяют на термофильные (политермные) – теплолюбивые и термофобные (олиготермные) – холодолюбивые.
- Солнечная радиация. Свет оказывает влияние на существование дневных и ночных форм животных. Существенное влияние оказывает длина светового дня. Явление фотопериодизма предопределяет развитие строго определенного количества поколений в течение года у насекомых, расширение ареалов в другие широтные пояса и др.
- Характер субстрата. Существует зависимость распространения животных от солености почвы. Наиболее чувствительны к засолению членистоногие. К примеру, жуки рода Bledius встречаются только на засоленных почвах. Химизм почвы может оказывать на животных опосредованное влияние, через кормовые растения.
- Растительный покров обуславливает характер биоценоза и выступает его индикатором. Например, в хвойных лесах Севера, где произрастают багульник, брусника, зеленые мхи можно встретить синицу-московку, глухаря, клестов, кедровку, соболя, бурундука, рысь.
Или искусственные: пруд, водохранилище, канал (рис. 4-6).

Рис. 5. Водохранилище ()

Каким бы ни был водоем, естественным, искусственным, он украшает нашу землю, радует нас своей красотой. В пресных водоемах мы берем воду, без которой не обойтись ни в быту, ни на производстве. В водоемах мы купаемся, рядом с ними загораем, по воде путешествуем на судах, перевозим грузы. Значение водоемов в природе велико. Пресная вода является важнейшим условием существования человека на Земле, а для животных, которые живут в воде, это еще и единственный дом. В воде есть все необходимое для жизни: свет, тепло, воздух и растворенные минеральные вещества.
Какие же растения растут и какие животные обитают в пресных водоемах? Оказавшись у водоема в теплое время года, вы могли наблюдать только тех его обитателей, которые живут на поверхности. Но жизнь в водоеме есть везде: и у берегов, и на поверхности, и в толще воды, у самого дна и на дне. На берегах водоемов можно увидеть листья и стебли камыша, тростника, рогозы, стрелолиста. Небольшая глубина позволяет этим растениям прикрепиться ко дну водоема. На значительно большей глубине растут кувшинка белая, кубышка желтая (рис. 7, 8). На гладкой поверхности воды плавают их цветки и широкие листья.

Рис. 7. Кувшинка белая ()

Рис. 8. Кубышка желтая ()
Как этим растения удалось приспособиться к жизни в сильно увлажненной почве, где почти нет кислорода? Если рассмотреть срез стеблей тростника, камыша, рогоза, то можно увидеть воздушные каналы, которые проходят в стеблях этих растений (рис. 9, 10).

Рис. 9. Тростник ()

Воздухоносные каналы есть и в листьях, и в корнях водных растений. У кувшинки белой и кубышки желтой черешки листьев и цветоножки, на которых сидят цветки, тоже пронизаны воздухоносными каналами, по которым проникает кислород, необходимый для дыхания. Срывая цветок, человек наносит вред всему растению. В место разрыва в растение начинает проникать вода, это приводит к загниванию подводной части и, в конечном счете, гибели всего растения.
Трава ряска в виде мелких зеленых пластинок тоже плавает на поверхности водоема, но корнями ко дну не прикрепляется, а в толще воды - мельчайшие зеленые водоросли, рассмотреть их можно только под микроскопом. Но их присутствие выдает цвет воды. Когда их в водоеме очень много, цвет воды становится зеленым.
Какую роль играют растения в жизни многочисленных жителей водоемов? Во-первых, зеленые растения под воздействием солнечного света из воздуха берут углекислый газ, а выделяют в воду кислород, необходимый для дыхания всех животных. Во-вторых, в зарослях водоема находят убежище и пищу птицы, земноводные, насекомые и их личинки, рыбы. Животные в водоемах есть везде: на поверхности и в толще воды, на берегу, на дне, на водных растениях. Главные связи между животными и растениями - пищевые. Вот водомерки (рис. 11) стремительно бегут по поверхности воды и охотятся на комаров и других мелких животных.

Рис. 11. Водомерка ()
Их длинные ножки снизу покрыты жиром, поэтому вода их и держит. А на водных растениях обитают улитки: прудовик и катушка (рис. 12, 13).

Рис. 12. Прудовик ()

Без кого же река жить не может? Очень маленькие рачки водоемов, дафнии и циклопы, живут и зимуют в воде. Их величина немного больше запятой в книжке (рис. 14, 15).
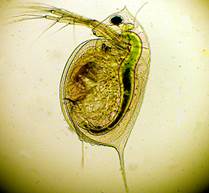
Самое примечательное у дафнии - длинные усы. Взмахнут усами, резко опустят, оттолкнутся от воды и подпрыгнут. У циклопа есть непарный лобный глазок, из-за которого он и получил свое название.

Без рачков река жить не может, так как они очищают воду от невидимых глазу бактерий, зеленых водорослей и мельчайших животных, если бы не рачки, река быстро бы переполнилась ими. Дафнии и циклопы, как и другие обитатели речки, питаются этими организмами, тем самым очищая воду. Сами служат кормом малькам рыб, моллюскам, головастикам, личинкам насекомых.
Неужели в реке кто-то без головы живет? Это моллюски, беззубки и перловицы (рис. 16).

Рис. 16 Моллюски ()
Сначала раковина, состоящая из двух продольных пластинок, будет лежать неподвижно, потом створки ее приоткроются и из нее высунется нога, головы ни у беззубки, ни у перловицы нет. Выдвинет беззубка ногу и воткнет ее в песок, раковина сдвинется с места. Переместится беззубка на 2-3 сантиметра, отдохнет - и снова в путь. Так и путешествует по дну реки. Пищу и воздух беззубка добывает прямо из воды. Приоткроет створки раковины и начинает втягивать воду, потом выбрасывает. В воде полно мельчайших животных, попадают они в раковину, так беззубка их специальными приспособлениями и задерживает. Дышит беззубка и ест, а заодно и воду очищает. И перловица также работает. Каждая за сутки очищает примерно 40-50 литров воды. Моллюсков, личинки насекомых, головастиков поедают рыбы, аисты, кулики, утки. Жук-плавунец охотится на других насекомых, а также червей, улиток, головастиков. Лягушки кормятся в прибрежных частях водоемов, в основном летающими насекомыми, а сами являются кормом для тритонов и хищных рыб, окуня и щуки. За рыбой и тритонами охотятся цапли, чайки, зимородки.
Основная пища рака - растительная. Но он охотно поедает животных, а также останки мертвых животных. Поэтому часто раков называют санитарами водоемов (рис. 17).

Раки за свою жизнь меняют панцирь. Органы чувств у речного рака развиты прекрасно, глаза выдвинуты вперед на тонких стебельках и состоят из огромного количества, 3000, крохотных глазков. Короткая пара усиков - органы обоняния, а длинные - осязания. Если хищник хватает рака за клешню, то рак ее обламывает и скрывается в норе. Потерянная клешня отрастет снова. Раки очень чувствительны к загрязнению воды, поэтому в местах, где они водятся, говорят об экологической чистоте водоемов.
У реки можно увидеть разных стрекоз: красотку, стрелку, лютку, они живут у реки постоянно (рис. 18).

Рис. 18. Стрекоза ()
Всем стрекозам нужна вода, потому что только там могут жить их личинки. Личинки не похожи на взрослых стрекоз, только глаза у них одинаковые. Каждый глаз состоит из почти 30 000 крохотных глазков (рис. 19).

Рис. 19. Личинка стрекозы ()
Оба глаза выпуклые, благодаря чему стрекоза одновременно может смотреть во все стороны (рис. 20). Все стрекозы - хищники, они охотятся в воздухе, на лету хватают насекомых.

Рис. 20. Глаза стрекозы ()
Личинка стрекозы, подкараулив добычу, выбрасывает вперед сильно удлиненную нижнюю губу. Обычно губы сложена и прикрывает голову, как маска. Личинка в большой мускульный мешок внутри тела всасывает воду, а потом силой выбрасывает. Получается водяной выстрел. Через год, а некоторые через 3, личинки выбираются на поверхность, шкурка личинки лопается, и из нее появляется стрекоза. Посидит несколько часов, расправит крылышки и улетает.
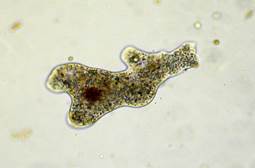
Кто же живет в капле воды? Если посмотреть в микроскоп, то откроется удивительный мир необычных существ. Вот почти прозрачный комочек, который все время меняется, - это амеба (рис. 21).
Другие существа напоминают крохотные туфельки, так они и называются. Тело у туфельки покрыто ресничками, каждая умело управляет этими ресничками и быстро плывет (рис. 22).
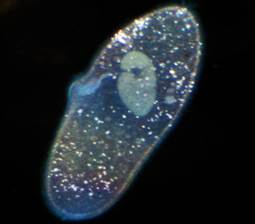
Рис. 22. Туфелька ()
Трубачи - самые красивые обитатели капли, голубые, зеленые, похожи на цветки вьюнков (рис. 23).
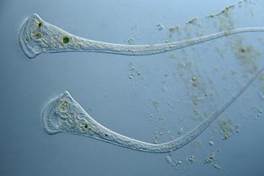
Двигаются трубачи неторопливо и только вперед. Если их что-то напугает, то они съеживаются и напоминают шарики. Амебы, туфельки и трубачи - одноклеточные организмы, питаются бактериями.
В капле воды также обитают и хищники. Это дидиний (рис. 24).
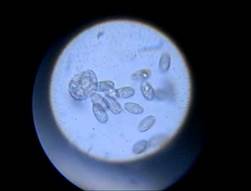
Хоть он и меньше туфельки, он не только смело нападает на нее, но и целиком заглатывает, раздуваясь как шар.
В пресном водоеме совместно обитают растения, животные, бактерии, все они хорошо приспособлены к жизни в воде и связаны между собой пищевыми цепочками. Когда растения и животные погибают, то они скапливаются на дне водоемов, под действием бактерий разрушаются и превращаются в соли, которые растворяются в воде и используются другими животными. Водоем - это природное сообщество.
Сегодня на уроке вы получили новое представление о пресном водоеме как о пресном сообществе и познакомились с его обитателями.
Список литературы
- Вахрушев А.А., Данилов Д.Д. Окружающий мир 3. - М.: Баллас.
- Дмитриева Н.Я., Казаков А.Н. Окружающий мир 3. - М.: ИД «Федоров».
- Плешаков А.А. Окружающий мир 3. - М.: Просвещение.
- Makuha.ru ().
- Youtube.com ().
- Sbio.info ().
Домашнее задание
- Какие пресные водоемы вы знаете?
- Каких животных можно встретить в водоемах?
- Почему говорят, что водоем - это природное сообщество?
Бурые и красные водоросли, общая характеристика, морфология, основы физиологии, специфика жизненных циклов, систематика, роль в биосфере и в жизни человека.
Красные водоросли, или Багрянки. Подавляющее большинство красных водорослей - морские обитатели, но встречаются они и в пресных водах.
Багрянки - многоклеточные организмы, имеющие нитчатую и пластинчатую структуру, и только немногие, наиболее примитивные, являются одноклеточными или колониальными формами. Многие из них - крупные растения, достигающие в длину нескольких десятков сантиметров, но среди них немало микроскопических форм. По форме красные водоросли бывают в виде нитей, кустиков, пластинок, корок, кораллов и др. Органами прикрепления служат ризоиды, присоски, подошвы. Окраска их слоевища меняется от ярко-красной до голубовато-зеленой и желтой. Это обусловлено тем, что в пластинчатых хроматофорах кроме хлорофилла содержатся красный пигмент фикоэритрин и синий фикоцианин.
Размножаются багрянки бесполым (спорами) и половым путем.
Бурые водоросли. Общим внешним признаком бурых водорослей, обитающих во всех морях земного шара, служит желтовато-бурая окраска их слоевища, обусловленная наличием в их клетках желтых и бурых пигментов (каротинов и ксантофиллов).
Слоевище бурых водорослей напоминает ветвистые кустики, пластинки, ленты, сложно расчлененные на стебле- и листовидные органы. Размеры таллома изменяются от нескольких сантиметров до 60- 100 м. Таллом нарастает в результате интеркалярного (вставочного) роста или вследствие постоянного деления верхушечной клетки. Для прикрепления кгрунту служат ризоиды или базальный диск - дисковидное разрастание в основании слоевища.
По морфологической и анатомической дифференцировке таллома бурые водоросли стоят на более высоком уровне, чем все другие группы. Среди них неизвестны ни одноклеточные, ни колониальные формы, ни талломы в виде простой неразветвленной нити. У большинства представителей талломы ложно- или истинно тканевого строения (выделяют ассимиляционные, запасающие, проводящие ткани).
Размножаются бурые водоросли бесполым (участками таллома или спорами) и половым способом. При половом размножении зигота без периода покоя прорастает в новое растение.
В цикле развития большинства бурых и красных водорослей происходит закономерное чередование поколений - гаметофита и спорофита.
Заросли крупных бурых водорослей иногда тянутся на десятки километров, образуя своеобразные подводные леса и луга на глубине 40- 100 м и более. Они служат кормом, местом размножения и укрытия многих видов животных, субстратом для микро-и макроорганизмов, одним из основных источников органики в водоемах.
Значение водорослей. Повсеместное распространение водорослей определяет их огромное значение в биосфере и хозяйственной деятельности человека. Благодаря способности к фотосинтезу они являются основными продуцентами громадного количества органических веществ в водоемах, которые широко используются животными и человеком.
Поглощая из воды углекислый газ, водоросли насыщают ее кислородом, необходимым всем живым организмам. Велика их роль в биологическом круговороте веществ.
В историческом и геологическом прошлом водоросли принимали участие в образовании горных и меловых пород, известняков, рифов, особых разновидностей угля, ряда горючих сланцев. Они явились родоначальниками растений, заселивших сушу.
Водоросли широко используются в народном хозяйстве, в том числе в пищевой, фармацевтической и парфюмерной промышленности. Их возделывают в больших масштабах в установках под открытым небом с целью получения биомассы как дополнительного источника белка, витаминов и биостимуляторов для животноводства. Так, установлено, что в морских водорослях присутствуют витамины А, В1 ,В2, В12, С и D, соединенияиода, брома и т. д.
Многие водоросли используются в пищу человеком. Наибольшее признание получила морская капуста (некоторые виды ламинарии и порфиры), которая, кроме того, применяется как лечебное и профилактическое средство при желудочно-кишечных расстройствах, склерозе, зобе, рахите и других заболеваниях.
Водоросли служат сырьем для получения многих ценных веществ: спиртов, лака, аммиака, органических кислот, иода, брома (бурые); агар-агара (красные). Агар-агар нашел широкое применение в биотехнологии в качестве твердой среды для культивирования бактерии, водорослей, грибов. В больших количествах его используют для изготовления мармелада, пастилы, мороженого и т, д.
В сельском хозяйстве водоросли применяются как органические удобрения под некоторые культуры и в качестве кормовой добавки в пищевом рационе домашних животных.
Некоторые водоросли (например, хлорелла, сценедесмус и др.) способны накапливать радионуклиды, что может использоваться для дополнительной очистки слабоактивных сточных вод АЭС.
Зеленые водоросли. Общая характеристика, морфология, основы физиологии, специфика жизненных циклов. Эволюция структур и способов размножения. Систематика, основные представители. Зеленые водоросли как предки высших растений. Роль в биосфере и в жизни человека.
Зеленые водоросли - самый обширный из всех отделов водорослей, насчитывающий по разным оценкам от 4 до 13 - 20 тысяч видов. Все они имеют зеленый цвет слоевищ, что обусловлено преобладанием в хлоропластах хлорофилла a и b над другими пигментами. Клетки некоторых представителей зеленых водорослей (Хламидомонас, Трентеполия, Гематококкус) окрашены в красный или оранжевый цвета, что связано с накоплением вне хлоропласта каротиноидных пигментов и их производных.
В морфологическом отношении они отличаются большим разнообразием. Среди зеленых водорослей встречаются одноклеточные, колониальные, многоклеточные и неклеточные представители, активно подвижные и неподвижные, прикрепленные и свободноживущие. Чрезвычайно велик и диапазон их размеров - от нескольких микрометров (что сравнимо по размерам с бактериальными клетками) до 1–2 метров.
Клетки одноядерные или многоядерные, с одним или несколькими хроматофорами, содержащими хлорофилл и каротиноиды. Хлоропласты покрыты двумя мембранами и обычно имеют стигму, или глазок, - фильтр, проводящий синий и зеленый свет к фоторецептору. Глазок состоит из нескольких рядов липидных глобул. Тилакоиды - структуры, где локализованы фотосинтетические пигменты - собраны в стопки (ламеллы) по 2–6. В переходной зоне жгутиков есть звездчатое образование. Жгутиков чаще всего два. Основной компонент клеточной стенки – целлюлоза.
У хлорофит встречаются различные типы питания: фототрофное, миксотрофное и гетеротрофное. Запасной полисахарид зеленых водорослей – крахмал – откладывается внутри хлоропласта. Хлорофиты также могут накапливать липиды, которые откладываются в виде капель в строме хлоропласта и в цитоплазме.
Многоклеточные слоевища нитевидные, трубчатые, пластинчатые, кустистые или иного строения и разнообразной формы. Из известных типов организации таллома у зеленых водорослей отсутствует только амебоидный.
Они широко распространены в пресных и морских водах, в почве и в наземных местообитаниях (на почве, скалах, коре деревьев, стенах домов и пр.). В морях распространено около 1/10 от общего количества видов, которые растут обычно в верхних слоях воды до 20 м. Среди них есть планктонные, перифитонные и бентосные формы. Иначе говоря, зеленые водоросли освоили три основных среды обитания живых организмов: воду – землю – воздух.
Зеленые водоросли обладают положительным (движение к источнику света) и отрицательным (движение от яркого источника света) фототаксисом. Кроме интенсивности освещения, на фототаксис влияет температура. Положительным фототаксисом при температуре 160°С обладают зооспоры видов родов Гематококкус, Улотрикс, Ульва, а также отдельные виды десмидиевых водорослей, у которых движение клеток осуществляется за счет выделения слизи через поры в оболочке.
Размножение. Для зеленых водорослей характерно наличие всех известных способов размножения: вегетативное, бесполое и половое.
Вегетативное размножение у одноклеточных форм происходит делением клетки пополам. Колониальные и многоклеточные формы хлорофит размножаются частями тела (слоевища, или таллома).
Бесполое размножение у зеленых водорослей представлено широко. Осуществляется чаще подвижными зооспорами, реже неподвижными апланоспорами и гипноспорами. Клетки, в которых образуются споры (спорангии), в большинстве случаев ничем не отличаются от остальных вегетативных клеток таллома, реже они имеют иную форму и более крупные размеры. Формирующиеся зооспоры могут быть голыми или покрытыми жесткой клеточной стенкой. Количество жгутиков у зооспор варьирует от 2 до 120. Зооспоры разнообразной формы: шаровидные, эллипсоидные или грушевидные, одноядерные, лишенные обособленной оболочки, с 2–4 жгутиками на переднем, более заостренном конце и хлоропластом в расширенном заднем конце. Обычно они имеют пульсирующие вакуоли и стигму. Зооспоры образуются одиночно или, чаще, в числе нескольких из внутреннего содержимого материнской клетки, выходят наружу через образующееся в оболочке круглое или щелевидное отверстие, реже вследствие ее общего ослизнения. В момент выхода из материнской клетки зооспоры иногда окружены тонким слизистым пузыре, в скором времени расплывающимся (род Улотрикс).
У многих видов вместо зооспор или наряду с ними образуются неподвижные споры – апланоспоры. Апланоспоры - споры бесполого размножения, у которых отсутствуют жгутики, но имеются сократительные вакуоли. Апланоспоры рассматривают как клетки, у которых приостановлено дальнейшее развитие в зооспоры. Они также возникают из протопласта клетки в числе одной или нескольких, но не вырабатывают жгутиков, а, приняв шаровидную форму, одеваются собственной оболочкой, в образовании которой оболочка материнской клетки не участвует. Апланоспоры освобождаются вследствие разрыва или ослизнения оболочек материнских клеток и прорастают после некоторого периода покоя. Апланоспоры с очень толстыми оболочками называются гипноспорами. Они обычно принимают на себя функцию покоящейся стадии. У автоспор, которые представляют собой уменьшенные копии неподвижных вегетативных клеток, отсутствуют сократительные вакуоли. Образование автоспор коррелирует с завоеванием наземных условий, в которых вода не может всегда присутствовать в достаточном количестве.
Половое размножение осуществляется гаметами, возникающих в неизмененных, слегка измененных или значительно преобразованных клетках – гаметангиях. Подвижные гаметы монадного строения, двужгутиковые. Половой процесс у зеленых водорослей представлен различными формами: гологамия, конъюгация, изогамия, гетерогамия, оогамия. При изогамии гаметы морфологически совершенно подобны друг другу и различия между ними являются чисто физиологическими. Зигота одевается толстой оболочкой, нередко со скульптурными выростами, содержит большое количество запасных веществ и прорастает сразу или после некоторого периода покоя. При прорастании содержимое зиготы у большинства видов делится на четыре части, которые выходят из оболочки и прорастают в новые особи. Значительно реже гаметы развиваются в новый организм без слияния, сами по себе, без образования зиготы. Такое размножение называется партеногенезом, а споры, образующиеся из отдельных гамет, – партеноспорами.
При гетерогамии обе гаметы различаются между собой по величине и иногда по форме. Более крупные гаметы, часто менее подвижные, принято считать женскими, меньшие по величине и более подвижные – мужскими. В одних случаях различия эти невелики, и тогда говорят просто о гетерогамии, в других весьма значительны.
Если женская гамета неподвижна и напоминает больше яйцеклетку, то подвижная мужская становится сперматозоидом, а половой процесс получает название оогамии. Гаметангии, в которых возникают яйцеклетки, называются оогониями, от вегетативных клеток они отличаются как по форме, так и по величине. Гаметангии, в которых образуются сперматозоиды, называются антеридиями. Зигота, получившаяся в результате оплодотворения яйцеклетки сперматозоидом, формирует толстую оболочку и называется ооспорой.
При типичной оогамии яйцеклетки крупные, неподвижные и развиваются чаще всего по одной в оогонии, сперматозоиды мелкие, подвижные, образуются в антеридии в большом количестве. Оогонии и антеридии могут развиваться на одной особи, в этом случае водоросли однодомные; если они развиваются на разных особях – двудомные. Оплодотворенная яйцеклетка одевается толстой бурой оболочкой; нередко соседние с ней клетки дают короткие веточки, которые обрастают ооспору, оплетая ее однослойной корой.
Жизненные циклы. У большинства представителей зеленых водорослей жизненный цикл гаплобионтный с зиготической редукцией. У таких видов диплоидной стадией является только зигота – клетка, получающаяся в результате оплодотворения яйцеклетки сперматозоидом. Другой тип жизненного цикла – гаплодиплобионтный со спорической редукцией – встречается у Ульвовых, Кладофоровых и некоторых Трентеполиевых. Для этих водорослей характерно чередование диплоидного спорофита и гаплоидного гаметофита. Гаплодиплобионтный жизненный цикл с соматической редукцией известен только у Празиолы. Наличие диплобионтного жизненного цикла у Бриопсидовых и Дазикладиевых подвергается сомнению.
У некоторых Улотриксовых одна и та же особь может давать начало как зооспорам, так и гаметам. В других случаях зооспоры и гаметы образуются на разных особях, т.е. жизненный цикл водорослей включает в себя как половую (гаметофит), так и бесполую (спорофит) форму развития. Спорофит обычно диплоидный, т.е. имеет в клетках двойной набор хромосом, гаметофит гаплоидный, т.е. имеет одинарный набор хромосом. Это наблюдается в тех случаях, когда мейоз происходит при образовании спор (спорическая редукция) и часть жизненного цикла водоросли от зиготы до образования спор проходит в диплофазе, а часть от споры до образования гамет в гаплофазе. Такой цикл развития характерен для видов рода Ульва.
В пределах Улотриксовых водорослей широко распространена зиготическая редукция, когда мейоз происходит при прорастании зиготы. Диплоидной в этом случае оказывается только зигота, весь остальной жизненный цикл протекает в гаплофазе. Значительно реже встречается гаметическая редукция, когда мейоз происходит при образовании гамет. В этом случае гаплоидными являются только гаметы, а весь остальной цикл диплоидный.
Систематика . До сих пор отсутствует единая устоявшаяся система зеленых водорослей, особенно в отношении группировки порядков в различные предлагаемые классы. Очень долго типу дифференциации таллома придавали основное значение при выделении порядков у зеленых водорослей. Однако в последнее время в связи с накоплением данных об ультраструктурных особенностях жгутиковых клеток, типе митоза и цитокинеза и др. очевидна гетерогенность многих таких порядков.
Отдел включает 5 классов: Ульвофициевые – Ulvophyceae, Брипсодовые – Bryopsidophyceae, Хлорофициевые – Chlorophyceae , Требуксиевые – Trebouxiophyceae , Празиновые – Prasinophyceae .
Экология и значение. Зеленые водоросли широко распространены по всему миру. Большинство из них можно встретить в пресных водоемах, но немало солоноватоводных и морских форм. Нитчатые зеленые водоросли, прикрепленные или неприкрепленные, наряду с диатомовыми и синезелеными являются преобладающими бентосными водорослями континентальных водоемов. Они встречаются в водоемах различной трофности (от дистрофных до эвтрофных) и с различным содержанием органических веществ (от ксено- до полисапробных), водородных ионов (от щелочных до кислых), при различных температурах (термо-, мезо- и криофилы).
Среди зеленых водорослей имеются планктонные, перифитонные и бентосные формы. В группе морского пикопланктона празиновая водоросль Остреококкус считается самой маленькой эукариотной свободноживущей клеткой. Есть виды зеленых водорослей, которые приспособились к жизни в почве и наземных местообитаниях. Их можно встретить на коре деревьев, скалах, различных постройках, на поверхности почв и в толще воздуха. В этих местообитаниях особенно распространены представители родов Трентеполия и Требуксия. Зеленые водоросли вегетируют в горячих источниках при температуре 35–52°С, а в отдельных случаях до 84°С и выше, нередко при повышенном содержании минеральных солей или органических веществ (сильно загрязненные горячие сточные воды заводов, фабрик, электростанций или атомных станций). Они также преобладают среди криофильных видов водорослей. Они могут вызывать зеленое, желтое, голубое, красное, коричневое, бурое или черное «цветение» снега или льда. Эти водоросли находятся в поверхностных слоях снега или льда и интенсивно размножаются в талой воде при температуре около 0 °С. Лишь немногие виды имеют стадии покоя, тогда как большинство лишены каких-либо специальных морфологических приспособлений к низким температурам.
В пересоленных водоемах преобладают одноклеточные подвижные зеленые водоросли – гипергалобы, клетки которых лишены оболочки и окружены лишь плазмалеммой. Эти водоросли отличаются повышенным содержанием хлористого натрия в протоплазме, высоким внутриклеточным осмотическим давлением, накоплением в клетках каротиноидов и глицерина, большой лабильностью ферментных систем и обменных процессов. В соленых водоемах они нередко развиваются в массовом количестве, вызывая красное или зеленое «цветение» соленых водоемов.
Микроскопические одноклеточные, колониальные и нитчатые формы зеленых водорослей приспособились к неблагоприятным условиям существования в воздушной среде. В зависимости от степени увлажнения их подразделяют на 2 группы: воздушные водоросли, обитающие в условиях только атмосферного увлажнения, и, следовательно, испытывающие постоянную смену влажности и высыхания; водновоздушные водоросли, подвергающиеся действию постоянного орошения водой (под брызгами водопада, прибоя и т. д.). Условия существования водорослей аэрофильных сообществ очень своеобразны и характеризуются, прежде всего, частой и резкой сменой двух факторов - влажности и температуры.
Сотни видов зеленых водорослей обитают в почвенном слое. Почва как биотоп имеет сходство и с водными и с воздушными местообитаниями: в ней есть воздух, но насыщенный водяными парами, что обеспечивает дыхание атмосферным воздухом без угрозы высыхания. Интенсивное развитие водорослей как фототрофных организмов возможно только в пределах проникновения света. В целинных почвах это поверхностный слой почвы толщиной до 1 см, в обрабатываемых почвах он немного толще. Однако в толще почвы, куда не проникает свет, жизнеспособные водоросли обнаруживаются на глубине до 2 м в целинных почвах и до 3 м – в пахотных. Это объясняется способностью некоторых водорослей переходить в темноте к гетеротрофному питанию. Многие водоросли сохраняются в почве в покоящемся состоянии.
Для поддержания своей жизнедеятельности почвенные водоросли имеют некоторые морфологические и физиологические особенности. Это относительно мелкие размеры почвенных видов, а также способность к обильному образованию слизи – слизистых колоний, чехлов и обверток. Благодаря наличию слизи, водоросли быстро поглощают воду при увлажнении и запасают ее, замедляя высыхание. Характерной чертой почвенных водорослей является «эфемерность» их вегетации – способность быстро переходить из состояния покоя к активной жизнедеятельности и наоборот. Они также способны переносить разные колебания температуры почвы. Диапазон выживаемости ряда видов лежит в пределах от -200 до +84 °С и выше. Наземные водоросли составляют важную часть растительности Антарктиды. Они окрашены почти в черный цвет, поэтому температура их тела оказывается выше температуры окружающей среды. Почвенные водоросли являются также важными компонентами биоценозов аридной (засушливой) зоны, где почва в летнее время нагревается до 60–80°С. Защитой от избыточной инсоляции служат темные слизистые чехлы вокруг клеток.
Своеобразную группу представляют эндолитофильные водоросли, связанные с известковым субстратом. Во-первых, это – сверлящие водоросли. Например, водоросли из рода Гомонтия сверлят раковины перловиц и беззубок, внедряются в известковый субстрат в пресных водоемах. Они делают известковый субстрат рыхлым, легко поддающимся различным воздействиям химических и физических факторов. Во-вторых, ряд водорослей в пресных и морских водоемах способны переводить растворенные в воде соли кальция в нерастворимые и отлагающие их на своих талломах. Ряд тропических зеленых водорослей, в частности Галимеда, откладывает в талломе карбонат кальция. Они принимают активное участие в постройке рифов. Гигантские залежи останков Галимеды, иногда достигающие 50 м в высоту, встречаются в континентальных шельфовых водах, связанных с Большим Барьерным Рифом в Австралии и других регионах, на глубине от 12 до 100 м.
Зеленые требуксиевые водоросли, вступая в симбиотические отношения с грибами, входят в состав лишайников. Около 85% лишайников содержат в качестве фотобионта одноклеточные и нитчатые зеленые водоросли, 10% - цианобактерии и 4% (и более) содержат одновременно синезеленые и зеленые водоросли. В качестве эндосимбионтов они существуют в клетках простейших, криптофитовых водорослей, гидр, губок и некоторых плоских червей. Даже хлоропласты отдельных сифоновых водорослей, например Кодиума, становятся симбионтами для голожаберных моллюсков. Эти животные питаются водорослями, хлоропласты которых остаются жизнеспособными в клетках дыхательной полости, причем на свету они очень эффективно фотосинтезируют. Ряд зеленых водорослей развивается на шерсти млекопитающих. Эндосимбионты, претерпевая морфологические изменения по сравнению со свободноживущими представителями, не теряют способности фотосинтезировать и размножаться внутри клеток хозяина.
Хозяйственное значение. Благодаря способности к фотосинтезу они являются основными продуцентами громадного количества органических веществ в водоемах, которые широко используются животными и человеком. Поглощая из воды углекислый газ, зеленые водоросли насыщают ее кислородом, необходимым всем живым организмам. Велика их роль в биологическом круговороте веществ . Быстрое размножение и очень высокая скорость ассимиляции (примерно в 3-5 раз выше, чем у наземных растений) приводят к тому, что за сутки масса водоросли увеличивается более чем в 10 раз. При этом в клетках хлореллы накапливаются углеводы (в селекционных штаммах их содержание достигает 60%), липиды (до 85%), витамины B, С и К. Белок хлореллы, на долю которого может приходиться до 50% сухой массы клетки, содержит все незаменимые аминокислоты. Уникальная способность видов Хлореллы ассимилировать от 10 до 18% световой энергии (против 1–2% у наземных растений) позволяет использовать эту зеленую водоросль для регенерации воздуха в замкнутых биологических системах жизнеобеспечения человека при длительных космических полетах и подводном плавании.
Ряд видов зеленых водорослей используют как индикаторные организмы в системе мониторинга водных экосистем. Наряду с фототрофным способом питания многие одноклеточные зеленые водоросли (хламидомонады) способны всасывать через оболочку растворенные в воде органические вещества, что способствует активному очищению загрязненных вод, в которых развиваются эти виды. Поэтому их применяют для очистки и доочистки загрязненных вод, а также как корм в рыбохозяйственных водоемах.
Некоторые виды зеленых водорослей используются населением ряда стран в пищу.
Поверхностные пленки зеленых водорослей имеют большое противоэрозионное значение. Слизистые вещества клеточных оболочек склеивают частицы почвы. Развитие водорослей влияет на структурирование мелкозема, придавая ему водостойкость и препятствуя выносу с поверхностного слоя.
Почвенные водоросли оказывают влияние и на рост и развитие высших растений. Выделяя физиологически активные вещества, они ускоряют рост проростков, особенно их корней.
Клетки водорослей способны аккумулировать из воды различные химические элементы, причем коэффициенты их накопления достаточно высоки. Мощными концентраторами являются пресноводные зеленые водоросли, особенно нитчатые. При этом интенсивность накопления в них металлов гораздо выше, чем в других пресноводных гидробионтах. Немалый интерес представляет способность водорослей концентрировать в себе радиоактивные элементы. Отмершие клетки водорослей удерживают накопленные элементы не менее прочно, чем живые, а в некоторых случаях десорбция из мертвых клеток меньше, чем из живых. Способность ряда родов (Хлорелла, Сценедесмус и др.) концентрировать и прочно удерживать в своих клетках химические элементы и радионуклиды позволяет использовать их в специализированных системах очистки для дезактивации промышленных сточных вод, например для дополнительной очистки слабоактивных сточных вод АЭС.
Некоторые зеленые водоросли являются антагонистами вируса гриппа, полиовируса и др. Выделяемые водорослями биологически активные вещества играют важную роль в обеззараживании воды и подавлении жизнедеятельности патогенной микрофлоры.
В специальных биологических прудах сообщества водорослей и бактерий используют для разложения и детоксикации гербицидов. Доказана способность ряда зеленых водорослей гидролизовать гербицид пропанил, который быстрее разрушается бактериями.
Тканевая дифференцировка высших растений, ее возникновение в эволюции. Типы тканей.
Среди растений, обитающих на Земле, есть одноклеточные, тело которых состоит из одной клетки. Это водоросли хлорелла, хламидомонада. У этих растений клетка представляет собой отдельный целостный организм. В ней протекают все процессы жизнедеятельности: питание, дыхание, образование и выделение веществ, размножение.
Растения, тело которых состоит из большого числа клеток, называются многоклеточными. Таких рас-тений большинство. Это некоторые водоросли, мхи, папоротники, голосеменные и цветковые растения. У многоклеточных растений клетки, как правило, различаются по строению и функциям. Одни выполняют функцию питания, другие - размножения, третьи - передвижения веществ.
Строение растений в процессе исторического развития постепенно усложнялось, увеличивалось чис-ло разных типов клеток. Если у одноклеточных водорослей тело состоит из одной клетки, то у мхов уже насчитывается около 20 разных типов клеток, у папоротников - около 40, а у покрытосеменных - около 80.
Группы клеток, сходные по строению, происхождению и приспособленные к выполнению одной или нескольких функций, называют тканью. Ткани, состоящие из одного типа клеток, называются простыми, а состоящие из разных типов – сложными или комплексными.
У растений различают образовательную ткань, покровную, механическую, проводящую, фотосинте-зирующую и запасающую.
Образовательная ткань (меристема). Эта ткань состоит из более или менее одинаковых клеток с тонкими оболочками, способных делиться. Клетки плотно прилегают друг к другу, содержат ядро, цитоплазму, не имеют заметных вакуолей. Они расположены в кончиках корней и верхушках побегов, у основания молодых листьев, стеблевых междоузлий, под корой стволов деревьев (камбий). Клетки образовательной ткани постоянно делятся, благодаря чему побег и корень растут в длину и в толщину, распускаются почки, бутоны, из семян вырастают проростки. Образовательная ткань обеспечивает рост растения, образование новых тканей и органов.
Покровная ткань. Клетки этой ткани плотно прилегают друг к другу и защищают органы расте-ний от неблагоприятных воздействий внешней среды: высыхания, механических повреждений. Благодаря покровной ткани растение взаимодействует со средой, через нее в клетку поступают необходимые вещества из окружающей среды и выделяются отработанные продукты. Например, через покровную ткань (устьица) листа в растении осуществляется газообмен и происходит испарение воды.
Механическая ткань. Образована эта ткань клетками с толстыми, нередко очень прочными оболочками, которые часто пропитываются жироподобными веществами. Поэтому ткань придает растению постоянную форму, обеспечивает его сопротивление на излом и изгиб. Механические ткани образуют скелет растения и обеспечивают его прочность, за что их называют еще опорными.
Фотосинтезирующая (ассимиляционная) ткань. Эта ткань состоит из тонкостенных живых клеток, в цитоплазме которых содержатся многочисленные хлоропласты. В них образуются органические вещества в процессе фотосинтеза. Фотосинтезирующая ткань имеет зеленую окраску. Она обусловлена со-держанием в хлоропластах хлорофилла - пигмента зеленого цвета. Кроме хлорофилла в клетках фотосинте-зирующей ткани в небольшом количестве содержатся и другие пигменты (желтые, оранжевые). Они погло-щают коротковолновые лучи и выполняют роль экрана, который защищает хлорофилл и клетку от губи-тельного воздействия этих лучей.
Запасающая ткань. Образована крупными живыми клетками с тонкими оболочками. В них откла-дываются запасные вещества: белки, жиры и углеводы. У одних растений запасные вещества отклады-ваются в семенах, у других - в корнях, стеблях, клубнях и т.д. В стеблях кактусов, в листьях алоэ хорошо развита водозадерживающая ткань, что позволяет растениям выдерживать солнечную жару, а также недостаток воды в почве. По мере расходования воды ткани растения теряют упругость, но после поглощения воды они восстанавливают свой прежний вид и размеры.
Проводящие ткани пронизывают все тело растения, образуя непрерывную разветвленную систе-му - от мельчайших корешков до самых молодых листьев. Проводящие ткани способствуют образованию восходящего и нисходящего тока в растении. Восходящий ток - это ток минеральных солей, растворенных в воде, идущих от корней по стеблю к листьям. Восходящий ток осуществляется по сосудам (трахеям) и тра-хеидам ксилемы. Нисходящий ток - это ток органических веществ, направляющихся от листьев к корням по ситовидным элементам флоэмы. Наиболее древними проводящими элементами ксилемы являются трахеиды - вытянутые клетки с за-остренными концами. Трахеиды имеют одревесневшую клеточную стенку с различной степенью утолще-ния. Различают кольчатые, спиральные, точечные, пористые и другие трахеиды. Сосуды представляют со-бой длинные полые трубки, образованные одним рядом мертвых клеток, поперечные перегородки между которыми растворились. Ситовидные трубки флоэмы - это живые клетки, вытянутые в длину, с ситовидными отвер-стиями на концах (в местах соприкосновения клеток). Флоэма и ксилема образуют сосудисто-волокнистые пучки. Флоэма и ксилема - комплексные ткани. Так, ксилема включает сосуды, трахеиды, паренхиму и древесинные волокна (не всегда). Флоэма состоит из ситовидных трубок и клеток-спутниц, паренхимы и иногда лубяных волокон.
Типы размножения высших растений.
I. Воспроизведение, сопровождающееся размножением: А - материнская особь, А -дочерние, подобные материнской.
II. Воспроизведение, не сопровождающееся размножением: Ж - материнская особь, перестающая существовать после образования дочерней.
III. Размножение без воспроизведения. Материнская особь А производит потомков В, не подобных ей.
IV. Образование потомства, не являющееся воспроизведением (потомок В не подобен материнской особи А) и не сопровождающееся размножением.
Типы размножения растений . Новые особи могут образоваться от родительских бесполым путем, т. е. без участия гамет и полового процесса, и половым путем, когда образованию дочерних особей обязательно предшествует половой процесс. При бесполом размножении в широком смысле родительская особь или разделяется на более или менее равные части (деление одноклеточных водорослей, партикуляция у многолетних трав, см. гл. V и VII), или отделяет от себя мелкие зачатки дочерних особей, способные развиться в самостоятельные растения. Разделение вегетативного тела или отделение от него вегетативных же зачатков, например почек, клубеньков,- это вегетативное бесполое размножение. При этом генотип потомства остается в принципе неизменным и каждый потомок" полностью воспроизводит материнское растение. Иногда же в качестве зачатков отделяются специализированные клетки, называемые спорами (греч. спора - посев, сеяние). У высших растений образование спор связано с редукцией числа хромосом, поэтому вырастающие из спор дочерние растения не похожи на материнские (не воспроизводят их). Размножение спорами хотя и является бесполым, поскольку спора прорастает без всяких слияний с другими клетками, но оно связано у высших растений с половым размножением в закономерном цикле воспроизведения (см. ниже).
При половом размножении генотип дочерних особей может изменяться и обогащаться за счет различных рекомбинаций индивидуальных признаков родителей.
Общая характеристика вегетативного размножения. Вегетативное размножение - это увеличение числа особей данного вида или сорта посредством отделения жизнеспособных частей вегетативного тела растения. Каждая отделившаяся часть живет какое-то время самостоятельно и, как правило, образует новые органы, нередко недостающие (на отделившемся побеге образуются корни, на частях корня - побеги). Таким образом, при вегетативном размножении обычна и типична регенерация - восстановление целого из части. Впрочем, довольно часто все необходимые органы создаются у будущей самостоятельной особи еще до отделения ее от материнской (например, новые розеточные побеги с придаточными корнями на концах усов земляники).
Цикл развития мохообразных и папоротникообразных в связи с приспособлением к наземным условиям жизни.
Папоротники имеют характерный, ни с чем не сравнимый вид. Это древесное или травянистое растение. Оно имеет видоизмененный побег, к которому при помощи черешка крепятся псевдолистки, или вайи. Это – первый эволюционный шаг к образованию у растений настоящей листовой пластины. Вайи исполняют две функции: первая – фотосинтез, вторая – спорообразование.
В грунте растение закрепляется при помощи подземного стебля – корневища. От него отходит множество вегетативных корней. В стебле папоротника сформированы ткани – проводящие и паренхимные, дающие растению возможность потреблять больше минеральных веществ и воды, чем его более низкоорганизованные коллеги по планете.
Жизненный цикл папоротника состоит из двух фаз – спорофита и гаметофита с преобладанием первой фазы над второй. На нижней части вайи образуются гаплоидные споры. Со временем спорангий раскрывается, споры падают на землю и прорастают. Именно этот заросток несет гаметы, женские и мужские. Но яйцеклетки и сперматозоиды одного растения вызревают в разное время, поэтому автооплодотворения не происходит. Как и у мхов, для совершения оплодотворения папоротникам необходима сильно увлажненная среда.
Из оплодотворенной зиготы развивается особь – спорофит. Сначала она использует питательные вещества, которые есть в заростке, а когда он отмирает, начинает питаться самостоятельно.
Мхи – двудомные растения, то есть на верхушках мужских растений расположены органы, продуцирующие сперматозоиды, а на верхушках женских – продуценты яйцеклеток. Но каждое растение независимо от пола обладает стеблем и листками. Они мелкие и содержат хлорофилл. У многих мхов листки нижнего яруса становятся желто-бурыми из-за разрушения пигмента в условиях недостаточной освещенности.
Корней у мхов нет. К грунту они крепятся ризоидами – многоклеточными волосообразными отростками.
Мхи размножаются спорами, которые вызревают в спорангии спорофита. Спорофит мха представлен ножкой с коробочкой. Но живет он недолго и быстро усыхает. Подсохшая коробочка открывается, и из нее просыпаются споры. Из них вырастает растение с гаплоидным набором хромосом – многолетнее, зеленое; женская или мужская особь. В жизненном цикле мха преобладает гаметофит над спорофитом.
Строение и развитие женского и мужского гаметофита у голосеменных и покрытосеменных растений.
Женский гаметофит у всех голосеменных развивается полностью внутри мегаспорангия и не выходит даже частично наружу, т. е. не соприкасается непосредственно с воздушной средой. Доступ к женскому гаметофиту осуществляется только через микропиле. Таким образом внутри семязачатка создаются наиболее благоприятные условия защиты женского гаметофита от высыхания. В результате происходит постепенная редукция и упрощение женского гаметофита и архегониев, возникает возможность очень раннего формирования яйцеклетки, и у некоторых голосеменных (вельвичия и гнетум) образуются даже особые неотенические безархегониальные гаметофиты.
Голосеменные отличаются от папоротников также развитием мужского гаметофита, строением и способом прорастания микроспор. У папоротников, где развитие гаметофита происходит обычно лишь после высеивания спор, прорастание спор происходит через так называемый тетрадный рубец, расположенный на проксимальном полюсе споры. У голосеменных, где мужской гаметофит сильно упрощается и его развитие ускоряется, первые деления ядра микроспоры происходят уже внутри микроспорангия. В связи с ранним развитием мужского гаметофита и образованием гамет еще внутри оболочки споры возникает необходимость в приспособлении, посредством которого микроспора может изменять свой объем. Таким приспособлением оказывается борозда на дистальном полюсе микроспоры, впервые возникающая у некоторых семенных папоротников и характерная для огромного большинства голосеменных. Борозда служит не только для регулирования объема пыльцевого зерна. Она становится местом выхода из микроспоры гаустории (у низших групп) или пыльцевой трубки (у гнетовых и хвойных), также являющихся новообразованиями. Таким образом, у голосеменных, в отличие от папоротников, отверстие для выхода содержимого микроспоры образуется на дистальном полюсе. Гаустория (присоска) типа саговниковых растет горизонтально и служит лишь для прикрепления и питания мужского гаметофита; настоящая пыльцевая трубка хвойпых и гнетовых растет вертикально и служит главным образом для проведения спермиев к яйцеклеткам, т. е. является проводником (вектором), а не только присоской. Хотя обычно оба эти образования называют пыльцевыми трубками, но морфологически и функционально они очень различны.
Наконец, следует отметить характерное для голосеменных (за исключением вельвичии, гнетума и секвойи вечнозеленой) нуклеарное (ядерное) дробление зиготы. В этом отношении они отличаются не только от низших групп, но и от покрытосеменных, для которых (за исключением лишь рода пион) характерно клеточное дробление зиготы.
Циклы развития голосеменных представляют собой как бы переходную ступень между циклами развития папоротников и покрытосеменных. Так, например, сосна образует шишки двоякого рода - мелкие мужские, не более 2,5 см в длину, и крупные женские, достигающие у некоторых видов 45 см в длину. Женская шишка состоит из ряда чешуек, на поверхности каждой из которых находится по две семяпочки. В каждой семяпочке имеется диплоидная макроспоровая материнская клетка. Последняя делится мейотически, с образованием четырех гаплоидных макроспор, из которых лишь одна функционирует и развивается в многоклеточный макрогаметофит. На каждом таком макрогаметофите имеется по 2- 3 женских половых органа (архегония), содержащих по одной крупной яйцеклетке. На нижней стороне каждой чешуйки мужской шишки находится по два микроспорангия. В этих микроспорангиях имеется большое число микроспоровых материнских клеток, каждая из которых делится мейотически и образует четыре микроспоры. Находящиеся в микроспорангии, или в пыльцевом мешочке, микроспоры подвергаются делению и образуют четырехклеточный микрогаметофит, или пыльцевое зерно. Свободная пыльца разносится ветром. Достигнув женской шишки, пыльцевое зерно проникает в семяпочку через особое отверстие - микропиле - и соприкасается с макроспорангием.
Может пройти больше года, прежде чем одна из клеток пыльцевого зерна превратится в пыльцевую трубку, которая прорастает через макроспорангий и достигает макрогаметофита. Другая клетка пыльцевого зерна делится, образуя не подвижный сперматозоид, как у низших растений, а два мужских генеративных ядра. Когда конец пыльцевой трубки достигает шейки архегония и вскрывается, из него выходят два мужских ядра, которые оказываются рядом с яйцеклеткой. Одно из них сливается с ядром яйцеклетки, образуя диплоидную зиготу, а другое исчезает. После оплодотворения зигота делится и дифференцируется, образуя зародыш спорофита, окруженный тканями женского гаметофита, а также тканями материнского спорофита. Весь этот комплекс представляет собой семя.
Ткани макрогаметофита, снабжающие развивающийся зародыш питательными веществами, образуют эндосперм. Однако они состоят из гаплоидных клеток, а не из триплоидных (3n), как клетки эндосперма покрытосеменных, хотя и те и другие служат для обеспечения зародыша питанием. После кратковременного периода роста, в течение которого формируются несколько листовидных семядолей, эпикотиль (дающий начало стеблю) и гипокотиль (дающий начало первичным корням), зародыш переходит в состояние покоя и остается в этом состоянии, пока не упадет на землю. Попав в благоприятные условия, семя прорастает и развивается в зрелый спорофит - сосну.
Цикл развития покрытосеменных растений.
У покрытосеменных еще происходит смена поколений - спорофита и гаметофита, однако гаметофит редуцирован до нескольких клеток, находящихся в тканях цветка спорофита. Спорофит - это обычные, хорошо знакомые нам деревья, кустарники или травы. Не у всех растений цветки легко различимы; мелкие зеленые цветки злаков и некоторых деревьев существенно отличаются от тех ярко окрашенных образований, которые мы обычно называем цветками.
Цветок покрытосеменных - это измененный побег, содержащий вместо обычных зеленых листьев концентрически расположенные листья, модифицированные для выполнения функции размножения. Типичный цветок состоит из концентрически расположенных элементов четырех типов, прикрепленных к цветоложу - расширенному концу цветоносного стебля. Самые внешние элементы - чашелистики - обычно бывают зеленого цвета и больше всех похожи на настоящие листья. Внутри кольца чашелистиков расположены лепестки, в большинстве случаев ярко окрашенные для привлечения насекомых или птиц, обеспечивающих опыление.
Непосредственно внутри кольца лепестков находятся тычинки - мужские части цветка. Каждая тычинка состоит из тонкой тычиночной нити с расположенным на ее конце пыльником. Пыльник представляет собой группу пыльцевых мешков (микроспорангиев), каждый из которых содержит материнские клетки микроспор, так называемые материнские клетки пыльцы. В результате мейоза каждая из этих диплоидных клеток образует четыре гаплоидные микроспоры, которые после деления ядра превращаются в молодые микрогаметофиты, или пыльцевые зерна.
В самом центре цветка расположено кольцо пестиков (или один пестик, образовавшийся в результате слияния нескольких пестиков). Пестик состоит из утолщенной полой нижней части - завязи и отходящего от нее длинного тонкого столбика, который вверху заканчивается уплощенным рыльцем. Последнее обычно выделяет липкую жидкость для улавливания и удерживания попавших на пестик пыльцевых зерен. Все эти части цветка чрезвычайно разнообразны по числу, расположению и форме. Цветок, содержащий и тычинки и пестики, называется обоеполым; цветки, лишенные либо тычинок, либо пестиков, - однополыми. Однополые цветки, содержащие только тычинки, называются тычиночными; однополые цветки, содержащие только пестики, называются пестичными. Ива, тополь и финиковая пальма относятся к числу растений, у которых одни особи несут только тычиночные цветки, а другие - только пестичные. Органы размножения цветковых растений - тычинки, пестики, рыльце, столбик и т. д. - были изучены и получили свои названия до того, как стали известны различные стадии чередования поколений, и до того, как был выявлен параллелизм основных черт цикла развития мхов, папоротников и цветковых растений.
В завязи, расположенной у основания пестика, находятся одна или несколько семяпочек. Последние представляют собой макроспорангий, окруженный 1 или 2 интегументами. Как правило, каждая семяпочка содержит одну материнскую клетку макроспоры, которая в результате мейоза образует четыре гаплоидные макроспоры. Одна из макроспор развивается в макрогаметофит; остальные три разрушаются. Ход развития макрогаметофита специфичен для каждого вида; в типичном случае макроспора значительно увеличивается и ее ядро делится. Два дочерних ядра мигрируют на противоположные концы клетки, каждое из них делится, а затем делятся и эти дочерние ядра. Образующийся таким образом макрогаметофит, называемый зародышевым мешком, представляет собой восьмиядерную клетку с четырьмя ядрами у каждого конца. По одному ядру с каждого конца перемещается к центру клетки; эти два ядра, лежащие рядом в центре клетки, называются полярными ядрами. Одно из трех ядер, лежащих на одном конце макроспорофита, становится ядром яйцеклетки, а два других и три ядра, находящиеся на другом конце, исчезают.
Гаплоидная микроспора развивается внутри пыльцевого мешка в микрогаметофит, или пыльцевое зерно. Ядро микроспоры делится, образуя крупное ядро пыльцевой трубки и меньшее по размерам генеративное ядро. В большинстве случаев на этой стадии пыльца освобождается и переносится ветром, насекомыми или птицами на рыльце того же или ближайшего цветка. Попав на рыльце, пыльца прорастает. При прорастании пыльцы образуется пыльцевая трубка, растущая по столбику вниз к семяпочке. Кончик пыльцевой трубки выделяет ферменты, растворяющие клетки столбика, так что создается возможность для дальнейшего ее прорастания. Ядро трубки остается в кончике растущей пыльцевой трубки. Генеративное ядро мигрирует в пыльцевую трубку и делится с образованием двух ядер - ядер спермиев. Зрелый мужской гаметофит состоит из пыльцевого зерна и пыльцевой трубки, ядра трубки и двух ядер спермиев, а также некоторого количества связанной с ними цитоплазмы.
Проникнув в макрогаметофит через микропиле, кончик пыльцевой трубки лопается, и оба генеративных ядра проникают в макрогаметофит. Одно из этих ядер перемещается к ядру яйцеклетки и сливается с ним; образующаяся в результате диплоидная зигота дает начало новому поколению спорофита. Другое генеративное ядро перемещается к двум полярным ядрам, после чего все три ядра сливаются и образуют ядро эндосперма, содержащее тройной набор хромосом. Иногда два полярных ядра сливаются в одно еще до появления генеративного ядра. Описанное явление двойного оплодотворения, приводящее к возникновению диплоидной зиготы и триплоидного (с тройным набором хромосом) эндосперма, специфично и характерно для цветковых растений.
После оплодотворения зигота многократно делится и формирует многоклеточный зародыш. В результате делений ядра эндосперма образуются клетки эндосперма, наполненные пита тельными веществами. Эти клетки, окружающие зародыш, снабжают его питанием. После оплодотворения чашелистики, лепестки, тычинки, рыльце и столбик обычно завядают и опадают. Семяпочка вместе с содержащимся в ней зародышем превращается в семя; ее стенки утолщаются и превращаются в жесткие наружные покровы семени. Семя состоит из зародыша и эндосперма с запасом питательных веществ, заключенных в прочную оболочку, которая возникла из стенки семяпочки. Благодаря семенам вид имеет возможность расселяться в новые местообитания и переживать периоды неблагоприятных внешних условий (например, зимой), губительные для взрослых растений.
Плоды . Завязь - нижняя часть пестика, содержащая семяпочки, - разрастается и превращается в плод. Таким образом, число семян, содержащихся в плоде, соответствует числу семяпочек. В строго ботаническом смысле слова плод - это зрелая завязь, содержащая семена - зрелые семяпочки. В обыденной жизни мы называем плодами такие ароматные, мясистые образования, как виноград, ягоды, яблоки, персики, вишни. Но бобы фасоли и гороха, зерна кукурузы, помидоры, огурцы и дыни, а также орехи, репей и крылатые плоды клена - это тоже плоды. Настоящий плод развивается только из завязи. Плод, возникающий из чашелистиков, лепестков или цветоложа, называется ложным плодом. Плоды яблони состоят преимущественно из разросшегося мясистого цветоложа; только сердцевина яблока происходит из завязи.
Метаморфоз вегетативных органов.
Метаморфозы - видоизменения органов, возникшие в процессе эволюции растений как следствие изменения или смены функций органов под влиянием комплекса условий среды.
Корнеплоды и корневые шишки служат для запасания веществ, необходимых для образования семян. Корнеплоды образуются при утолщении главного корня и нижней части побега (свёкла, брюква, турнепс, репа, редис, редька, морковь, петрушка, вех, борщевик), а корневые шишки формируются на боковых корнях (георгины, аспарагус, зопник клубненосный, лабазник шестилепестный) двух - или многолетних растений.
Втягивающие корни могут укорачиваться у своего основания. За счёт этого они втягивают луковицы, корневища, клубни глубже в почву, где их почки защищены от неблагоприятных факторов. Втягивающие корни легко узнаются по поперечным полоскам на утолщённых основаниях. Они характерны для такого растения, как лилия - саранка, у которой с возрастом луковица погружается всё глубже в почву. Хорошо выражены у клубнелуковиц гладиолуса. Очевидно, их можно обнаружить и у других растений с луковицами, корневыми шишками и погружающимися корневищами. Известно, что у земляники, медуницы, копытня, фиалки, манжетки, гравилата надземные побеги с чешуевидными и зелёными розеточными листьями теряют фотосинтезирующие листья и втягиваются в почву придаточными корнями, становясь корневищами.
Бактериальные клубеньки. Боковые корни ряда растений приспособлены к симбиозу с некоторыми азотофиксирующими бактериями. В клубеньках из молекулярного азота воздуха синтезируются органические вещества, часть из которых используется растением. Кроме бобовых, способны образовывать на корнях клубеньки лох, ольха, облепиха. Бобовые, благодаря дополнительному источнику азота, богаты белками. Они дают ценные пищевые и кормовые продукты, обогащают почву азотистыми веществами (на 1 га почвы 200-300 кг азота) поэтому их часто используют в качестве "зелёных удобрений", а облепиху - при рекультивации нарушенных земель.
Микориза (грибокорень). Корни многих растений сожительствуют с почвенными грибами, образуя микоризу ("грибокорень"). Гифы грибов облегчают корням восприятие воды и минеральным веществ из почвы, передают им некоторые органические вещества витамины, регуляторы роста. Гриб получает от высших растений углеводы и другие питательные вещества. Микоризу образует большая часть дикорастущих и культурных трав и деревьев - представителей голосеменных, однодольных (75%) и двудольных (80-90% от общего числа видов). Растения, например древесные лучше растут в присутствии гриба, а семена орхидных только в этом случае прорастают. У деревьев грибные нити обычно располагаются снаружи (в виде чехла) и в поверхностных тканях корней, лишенных в этом случае корневых волосков. При сожительстве с травами споры грибов проникают, как правило, внутрь корней, которые могут усиленно при этом ветвиться и образовывать вздутия. Выпадение кислотных дождей вызывает сокращение видового разнообразия микоризообразующих грибов и разрушение микория Эти данные были получены при изучении сосны, берёзы и пихты! Видоизменения побегов. Побеги могут накапливать в паренхиме стебля или листьев различные вещества, необходимые для цветения или перенесения засушливых условий. Запасающие побеги обычно находятся под землёй и несут на себе чешуевидные листы и почки возобновления. Благодаря этому растения становятся многолетними, могут размножаться в условиях где затрудняется семеношение и захватывать новые территории. Иногда вместо листья функцию фотосинтеза выполняют стебли. Побеги могут превращаться в усики или колючки.
Отдел Зеленые водоросли - Chlorophyta.
Общее число видов около 15 тыс. Распространены повсеместно, преимущественно в пресных водоемах, некоторые в морях и очень немногие в условиях периодического увлажнения на почве, стволах деревьев, заборах, цветочных горшках и т. д.
На примере представителей этого отдела можно проследить два направления эволюции: от одноклеточных одноядерных форм до сифональных многоядерных, высшая ступень этой линии - каулерпа (род Caulerpa); от одноклеточных форм через колониальные к многоклеточным нитчатым и далее к многоклеточным с более или менее дифференцированным талломом, имитирующим по внешнему виду органы высших растений, высшая ступень этой линии - хара (род Сhara).
Органы движения у подвижных форм - два, реже четыре жгутика одинаковой длины и формы. Клетки одноядерные, но могут быть и многоядерными (сем. кладофоровые - Cladophoraceae). Хлоропласты в большинстве случаев с пиреноидами, разнообразные по форме, величине и по числу их в клетке. Пигменты - хлорофилл, каротиноиды. Запасные продукты - крахмал и масло. Размножение вегетативное, бесполое и половое. Половой процесс известен почти у всех видов и отличается большим разнообразием: изогамия, гетерогамия, оогамия, соматогамия (гологамия, конъюгация).
Зеленые водоросли делят на три класса: Равножгутиковые, Конъюгаты и Харовые.
Класс Равножгутиковые - Isocontophyceae.
Самый большой класс по числу видов. Таллом одноклеточный, колониальный, многоклеточный. В жизненном цикле имеется более или менее длительная подвижная фаза.
Хламидомонада (род. Chlamydomonas). Виды этого рода живут обычно в мелких загрязненных водоемах и лужах и часто вызывают «цветение» воды. Это одноклеточные водоросли разнообразной формы: округлой, овальной, яйцевидной. Стенка пектиново-целлюлозная. На переднем конце имеются два цитоплазматических жгутика. Хлоропласт чашевидный, вогнутой поверхностью обращен к переднему концу клетки. В базальной части хлоропласта имеется довольно крупный пиреноид, окруженный гранулами запасного крахмала, а в верхней части - стигма («глазок»). В цитоплазме, заполняющей углубление хлоропласта, находится ядро, а у основания жгутиков - пульсирующая вакуоль.
При благоприятных условиях хламидомонада размножается бесполым способом: протопласт делится митотически на два, четыре или восемь частей, из которых еще в материнской клетке формируются зооспоры, по общей структуре тождественные взрослым особям, но меньшего размера и без целлюлозной стенки. Вследствие ослизнения стенки материнской клетки они освобождаются, вырастают до размеров взрослых особей и строят новую клеточную стенку. При недостатке воды и кислорода хламидомонада сбрасывает.жгутики и выделяет слизь. При этом протопласт сохраняет способность к делению. С наступлением благоприятных условий вновь образовавшиеся клетки формируют жгутики, освобождаются от слизи и вырастают до обычных размеров. Половой процесс чаще изогамный, однако у некоторых видов отмечена гетерогамия и даже оогамия. Сформировавшаяся зигота наполняется, запасными продуктами и вырабатывает толстую стенку. Затем наступает период покоя. При благоприятных условиях содержимое зиготы делится путем мейоза, в результате формируются четыре гаплоидные зооспоры.
Хлорелла (род Chlorella). Виды этого рода широко распространены в пресных водоемах, морях, в почве, на корке стволов деревьев. Иногда входят в состав лишайников. Таллом одноклеточный. Клетка округлой формы, по структуре напоминает хламидомонаду, но без жгутиков и пульсирующей вакуоли. Споры не имеют жгутиков. Их называют апланоспорами. В материнской клетке образуются восемь спор, которые, разрастаясь, освобождаются и пассивно переносятся током воды. Бесполое размножение у хлореллы осуществляется очень быстро. Половой процесс отсутствует (по другим данным, у некоторых видов имеется). В клетках хлореллы накапливается много запасных продуктов, витаминов, антибиотиков, поэтому ее культивируют для использования с различными целями.
Улотрикс (род Ulothrix). Виды этого рода распространены в реках. Таллом нитчатый, неветвящийся, состоит из одного ряда одинаковых клеток, нарастает верхушкой, прикрепляется к субстрату бесцветной базальной клеткой. Хлоропласт имеет форму кольца или полукольца и занимает постенное положение. Ядро одно. При бесполом размножении в любой из клеток, кроме базальной, образуются четырехжгутиковые зооспоры. Половой процесс изогамный. Гаметы мелкие, двухжгутиковые, образуются также в любой из клеток. Сливаются только гаметы от разных особей (гетероталлизм). Зигота делится путем мейоза. В результате образуются четыре гаплоидные зооспоры, прорастающие во взрослые нити. Весь жизненный цикл проходит в гаплоидной фазе, диплоидна только зигота.
Каулерпа (род Caulerpa). Виды этого рода - морские водоросли, имеющие стелющийся по субстрату сифональный таллом, длиной до 50 см, а иногда и больше. Внешне он напоминает корневище с придаточными корнями и крупными листьями. Это как бы одна гигантская клетка с единым протопластом, с множеством ядер и хлоропластов. Полость таллома не имеет перегородок, но пересекается целлюлозными опорными тяжами. Собственно бесполого размножения нет, иногда бывает вегетативное размножение частями таллома. Половой процесс изогамный. Весь жизненный цикл проходит в диплоидной фазе. Мейоз осуществляется перед образованием изогамет.
Класс Конъюгаты - Conjugatophyceae.
Таллом многоклеточный нитчатый или одноклеточный без жгутиков. Половой процесс в виде соматогамии (конъюгации). Зооспор и гамет нет.
Спирогира (род Spirogyra). Многочисленные виды этого рода живут в пресных водоемах - в реках, прудах, озерах и торфяных болотах. Нитчатый таллом состоит из одного ряда клеток. Хлоропласты по 1 - 2 в клетке расположены в постенном слое цитоплазмы. Они имеют вид спирально закрученных лент с пиреноидами, края лент часто бывают зазубренные. Ядро находится в центре клетки и погружено в цитоплазму, тончайшие нити которой тянутся к ее постенному слою. Вакуолей несколько. Растет спирогира за счет деления клеток. Вегетативное размножение происходит обрывками таллома. Половой процесс осуществляется следующим образом: две гетероталличные особи располагаются параллельно; в их клетках возникают выпячивания стенок, растущие навстречу друг другу; в месте стыка стенки расслизняются, образуется конъюгационный канал, по которому протопласт из клетки одной особи, условно мужской, переходит в клетку женской особи. Половой процесс завершается образованием крупной шаровидной зиготы, которая вырабатывает толстую стенку и запасные продукты в виде масла. После периода покоя зигота делится мейозом. При этом образуются четыре гаплоидные клетки, три из них отмирают, а одна прорастает в новую особь. Таким образом, жизненный цикл проходит в гаплоидной фазе, диплоидна только зигота.
Класс Харовые - Charophyceae.
Крупные водоросли со сложно расчлененным талломом. Они живут чаще всего в водоемах с пресной водой (озерах, старицах рек), где образуют густые заросли. Бесполое размножение с помощью зооспор отсутствует. Вегетативное осуществляется специальными «клубеньками», образующимися на ризоидах, или частями таллома. Органы полового размножения - оогонии и антеридии - многоклеточные. Харовые - эволюционно наиболее продвинутые зеленые водоросли.
Хара (род Chara). У видов этого рода таллом достигает нескольких десятков сантиметров в длину. Он расчленен как бы на «узлы» и «междоузлия», от «узлов» отходят ответвления. Осевая часть таллома состоит из срединной крупной длинной клетки, которая окружена более мелкими. Длинные клетки вдоль таллома чередуются с более короткими. С помощью ризоидов таллом прикрепляется ко дну водоема.
Вегетативное размножение осуществляется «клубеньками», образующимися на ризоидах. При половом размножении в пазухах некоторых боковых одноклеточных ответвлений образуются оогонии и антеридии. Оогонии имеет продолговато-шаровидную форму. Стенка его состоит из спирально закрученных удлиненных клеток, заканчивающихся на вершине пятью короткими клетками (коронкой). Внутри находится яйцеклетка. Антеридии мельче оогониев и имеют шаровидную форму. В зрелом состоянии они окрашены в оранжевый цвет. Стенка антеридия состоит из восьми треугольных клеток - щитков. От каждого щитка внутрь отходит длинная клетка (рукоятка) с шаровидной клеткой на верхушке (головка), которая образует спермагенные нити. В клетках последних формируются сперматозоиды с двумя одинаковыми жгутиками. Оплодотворенная яйцеклетка разрастается в зиготу (ооспору), которая вступает в период покоя. Прорастанию ее предшествует мейоз. Затем образуется гаплоидная короткая неветвистая нить - предросток, из которой вырастает новое растение. Жизненный цикл проходит в гаплоидной фазе, диплоидна только зигота.
Лекция № 9
Высшие споровые растения.
Высшие растения.
У большинства высших растений тело дифференцировано на органы - корень, стебель и листья, состоящие из хорошо обособленных тканей. В жизненном цикле высших растений четко выражено чередование спорофита (2n) и гаметофита (n). Органы полового размножения многоклеточные. Женский - архегоний - состоит из расширенной нижней части - брюшка, где формируется яйцеклетка, и верхней суженной - шейки, раскрывающейся при созревании яйцеклетки. Мужской орган полового размножения - антеридий - имеет вид мешка, внутри которого образуется.множество сперматозоидов. У голосеменных антеридии подверглись редукции, а у покрытосеменных редуцированы и антеридии, и архегоний. Из зиготы у высших растений образуется зародыш - зачаток спорофита.
Отдел Моховидные - Bryophyta.
Общее число видов около 35 тыс.
Строение . В жизненном цикле моховидных, как и других высших растений, имеется чередование двух фаз: спорофита и гаметофита. Однако доминирует (преобладает) гаметофит, в то время как у всех других высших растений доминирует спорофит. Именно поэтому моховидные рассматривают как самостоятельную боковую ветвь в эволюции высших растений.
Моховидные по своей организации и экологии еще близки к водорослям. Как и у водорослей, у них нет сосудов и корней. Некоторые примитивные представители имеют вегетативное тело в виде стелющегося таллома с верхушечным (дихотомическим) ветвлением, похожего на таллом водорослей. Оплодотворение связано с водой. Среди моховидных, как и среди водорослей, нет одревесневающих форм.
Распространение . Моховидные распространены на всех континентах мира, но неравномерно. В тропических странах - преимущественно в горах. Незначительное число видов растет в засушливых условиях, например в степях. Некоторые виды ведут эпифитный образ жизни на корке деревьев или водный. Основное же разнообразие видов сосредоточено во влажных местах Северного полушария, в областях с умеренным и холодным климатом. В сложении растительного покрова, особенно тундр, болот и лесов, им принадлежит важная роль.
Классификация . Моховидные подразделяют на три класса: Антоцеротовые, Печеночники, Листостебельные мхи. Наибольшее значение имеют два последних класса.
Класс Печеночники - Hepaticopsida.
Общее число видов около 10 тыс. Распространелы повсеместно. Примитивность строения тела печеночников свидетельствует об их древности.
Маршанция обыкновенная (Marchantia polymorpha) - типичный представитель класса. Гаметофит в виде пластинчатого таллома, длиной 10 - 12 см, ветвление верхушечное. С обеих сторон он покрыт эпидермой. Верхняя эпидерма имеет вентиляционные отверстия - устьица. Они окружены специальными клетками, расположенными в четыре ряда. Под устьицами имеются воздушные камеры. Нижняя эпидерма дает выросты - одноклеточные ризоиды и красноватые или зеленоватые чешуйки, которые иногда принимают за редуцированные листья. Под верхней эпидермой расположена ассимиляционная ткань, состоящая из вертикальных столбиков паренхимных клеток с хлоропластами. Ниже находится слой тонкостенных бесхлорофилльных паренхимных клеток. Следовательно, таллом маршанции имеет дорсивентральное строение.
На верхней стороне таллома образуются особые веточки - подставки, а на них - органы полового размножения. Маршанция - двудомное растение. На одних экземплярах подставки имеют форму сидящей на ножке девятилучевои звездочки, между лучами которой на нижней стороне расположены архегонии. На других - подставки имеют форму сидящего на ножке восьмиугольного щитка, на верхней стороне которого расположены антеридии, погруженные в антеридиальные полости. В брюшке архегония образуется яйцеклетка. После ее слияния со сперматозоидом из зиготы образуется спорогон. Он представляет собой коробочку на короткой ножке, которая прикреплена к гаметофиту гаусторией. Внутри коробочки из спорогенных клеток в результате мейоза образуются гаплоидные споры, а также элатеры - мертвые удлиненные клетки со спирально утолщенной стенкой, служащие для разрыхления массы спор, а также для выбрасывания их из коробочки. В благоприятных условиях из споры развивается предросток, или протонема. Это небольшая нить. Из ее верхушечной клетки вырастает таллом маршанции.
Вегетативное размножение осуществляется выводковыми тельцами линзовидной формы, имеющими зеленый цвет. Они образуются на верхней стороне таллома в особых корзиночках в результате деления клеток, выстилающих их дно.
Виды маршанции имеют широкое распространение. Наиболее часто их можно встретить во влажных местах: на берегах озер и рек, по оврагам и в травянистом покрове под пологом леса.
Класс Листостебельные мхи - Bryopsida.
Общее число видов около 25 тыс. Многие виды распространены в приполярных странах Северного полушария. На огромных территориях в тундре, на болотах, в лесах они доминируют в растительном покрове, значительно влияя на обеспеченность суши влагой.
Гаметофит представляет собой прямостоячую стеблевидную ось - каулидий, покрытую листовидными выростами - филлидиями. Условно их можно называть стебель и листья. На нижней части стебля образуются многоклеточные ризоиды (не у всех). Ветвление боковое. Нарастание осей происходит в результате деления пирамидальной верхушечной клетки. Оно может быть моноподиальным или симподиальным. В соответствии с этим органы полового размножения и спорогон размещаются на вершине гаметофита или на боковых разветвлениях.
Класс подразделяют на три подкласса: Андреевые мхи, Сфагновые мхи, Бриевые (Зеленые) мхи. Наибольшее значение имеют два последних подкласса.
Подкласс Сфагновые мхи - Sphaqnidae.
Сфагновые мхи имеют довольно однообразное строение и поэтому трудны для определения. Гаметофит их представляет собой сильноветвящееся, особенно в верхней части, растение. Разветвления густо покрыты листьями. Сфагновые мхи живут в очень влажной среде. В связи с этим у них нет ризоидов и влага поступает непосредственно в стебель, который при основании с течением времени отмирает. Строение стебля несложное. В центре его находится сердцевина из тонкостенных паренхимных клеток, выполняющих проводящую и запасающую функции. Она окружена корой, состоящей из двух слоев: склеродермы, выполняющей механическую функцию, и гиалодермы, выполняющей водозапасающую функцию. Клетки гиалодермы крупные, мертвые, стенки их имеют круглые отверстия, через которые полости смежных клеток сообщаются друг с другом, а также с внешней средой. Иногда эти клетки несут спиральные утолщения. Лист состоит из одного ряда клеток, резко различающихся как по структуре, так и по выполняемой функции. Одни из них живые, хлорофиллоносные, другие - мертвые, относительно более крупные, со спирально утолщенными стенками, пронизанные отверстиями, по строению сходные с водозапасающими клетками гиалодермы, их называют гиалиновыми. Гиалиновые клетки способны накапливать и долго удерживать огромное количество воды, в 30 - 40 раз превышающее массу самого растения.
Гаметофиты - однодомные и двудомные. Антеридии образуются в пазухах листьев на разветвлениях стебля. Около них листья окрашены в красноватый цвет. Архегонии на укороченных разветвлениях. В результате слияния сперматозоида с яйцеклеткой возникает зигота, которая представляет собой начало диплоидной фазы - спорогона. Спорогон состоит из ножки и коробочки. Ножка сильно укорочена, луковицеобразная, но ко времени созревания спор верхушка стебля гаметофита сильно вырастает и выносит коробочку кверху (ложная ножка). В центре коробочки помещается округлая колонка, над которой размещен в виде свода спорангий со спорогенной тканью. Стенка коробочки прочная, многослойная. Наружный хлорофиллоносный слой содержит большое число недоразвитых устьиц. Коробочка имеет крышечку, которая во время созревания спор отскакивает, и споры рассеиваются. Элатер нет. Из спор образуется сначала зеленая пластинчатая протонема, а затем из почек, расположенных на ней, - взрослый гаметофит, который и доминирует в жизненном цикле.
Структура сфагновых отличается примитивностью: пластинчатая протонема, отсутствие проводящего пучка и ризоидов, слабая дифференциация коробочки.
Значение сфагновых в природе очень велико. Накапливая огромное количество воды и разрастаясь плотными дернинами, они вызывают заболачивание огромных пространств, достигающих зоны тундры. Для осушения их осуществляют агромелиоративные работы. С другой стороны, старые болота имеют важное хозяйственное значение для разработки залежей торфа. Нарастание пласта торфа в наиболее благоприятных условиях происходит медленно - слой толщиной 1 см образуется примерно за 10 лет.
Подкласс Бриевые (зеленые) мхи - Bryidae.
Число видов 24,6 тыс. Распространены более широко, чем сфагновые мхи. Живут в разнообразных экологических условиях от тундры и лесотундры до степей и пустынь. Наиболее типичные местообитания бриевых мхов, где они доминируют или формируют сплошной покров, - тундра, болота и некоторые типы лесов. Каждому местообитанию свойственны свои виды. Бриевые мхи по сравнению со сфагновыми отличаются большим разнообразием строения. Органы полового размножения закладываются у одних видов на главной оси, у других - на боковых. У некоторых видов ветвление не выражено.
Политрих обыкновенный, кукушкин лен (Polytrichum commune) - это один из распространенных представителей бриевых мхов. Он растет в лесу, на полянах, окраинах болот.
Стебель гаметофита прямостоячий, неветвистый, высотой 15 см и более, густо покрыт листьями. Подземная часть его простирается в почве почти горизонтально, на ней образуются ризоиды. В центре стебля находится концентрический проводящий пучок, состоящий из вытянутых клеток, сходных с трахеидами и ситовидными трубками. Он окружен паренхимой, также выполняющей проводящую функцию. С наружной стороны паренхима граничит со склеродермой (корой). Внешний слой ее, состоящий из бесцветных клеток, называют гиалодермой.
Листья расположены по спирали. Они состоят из линейной пластинки с заостренной зубчатой верхушкой и пленчатого влагалища. На верхней стороне листа расположены ассимиляционные пластинки. Жилка с механическими и проводящими элементами расширена.
Гаметофит двудомный. Архегонии бутылковидной формы расположены на верхушке женского гаметофита, антеридии мешковидной формы - на верхушке мужского. Между архегониями и антеридиями имеются стерильные нити - парафизы. После оплодотворения из зиготы образуется спорогон, состоящий из длинной ножки и коробочки. Коробочка прямостоячая или более или менее косо расположенная, призматическая, четырех-пятиграниая, покрыта ржаво-войлочным колпачком, образующимся из стенок архегония. Коробочка состоит из урночки и крышечки. Нижняя часть урночки сужена в шейку. На границе урночки и шейки в эпидерме имеются устьица. В центре урночки расположена колонка, которая у крышечки расширяется и формирует эпифрагму - тонкостенную перегородку, закрывающую урночку. Вокруг колонки расположен спорангий в виде цилиндрического мешка, прикрепленного к стенке и колонке особыми нитевидными образованиями. Урночка имеет специальное приспособление для рассеивания спор - перистом, представляющий собой ряд зубчиков с тупыми верхушками, расположенных по краю урночки. Между зубцами, способными к гигроскопическим движениям, и эпифрагмой имеются отверстия, через которые в сухую погоду высыпаются споры. Из споры вырастает протонема в виде зеленой ветвящейся нити. На ней формируются почки, из которых со временем развиваются взрослые гаметофиты.
Отделы Риниовидные - Rhyniophyta и Псилотовидные - Ps1lotophyta.
Отдел Риниовидные включает 2 - 3 рода только ископаемых растений. В жизненном цикле доминирует спорофит. Вегетативное тело его состоит из системы разветвленных теломов. Общая структура в надземной части телома очень своеобразна. Это еще не побег, поскольку листьев на осях телома нет. Главная ось хорошо выражена. Ветвление верхушечное (дихотомическое). В центре оси обособлена ксилема, окруженная флоэмой. Ксилема может быть расположена компактно в виде цилиндра или в виде лучей. Она состоит из трахеид. Периферическая (коровая) часть телома выполняет функцию фотосинтеза. В эпидерме находятся устьичные аппараты. На подземной части устьиц нет. Настоящих корней нет, их заменяют ризоиды. Спорангии расположены на верхушках телома, стенка спорангия многослойна. Гаметофиты риниовидных не найдены. Представитель - род риния (Rhynia), включающий два вида. Это травянистые растения высотой около 20 см, диаметром 3 мм. Подземная часть состоит из горизонтального телома, от которого перпендикулярно отходят воздушные оси.
К отделу Псилотовидные в современной.флоре относят два рода: псилот (Psilotum) и тмезиптер (Tmesipteris). Общее число видов 4 - 6. Оба рода имеют широкое распространение в тропической и субтропической зонах обоих полушарий.
Спорофит псилотовидных - эпифитное, реже наземное травянистое растение. Телом длиной 5 - 40 (до 100) см. Ветвление чаще верхушечное. Кора хорошо развита, выполняет функцию фотосинтеза. Устьичные аппараты примитивные. Листья мелкие, длиной 1 - 5 мм, шиловидные, плоские, без устьичных. аппаратов и жилок. Их можно рассматривать как выросты телома. Подземная часть представлена корневищем с ризоидами. Корней нет. Спорангии срастаются по 2 - 3 (синангии), вскрываются продольной щелью. Споры одинаковой величины. Структура спорофита псилотовидных свидетельствует о близости к риниовидным.
Гаметофит обоеполый, без хлорофилла, радиально симметричный, ветвление верхушечное. Длина его примерно 20 мм, диаметр 2 мм, Питается сапрофитно при помощи грибов, с которыми вступает в симбиоз. Поверхность покрыта ризоидами. Живет чаще под землей. Оплодотворение связано с водой.