Бесполое размножение грибов примеры. Способы распространения грибов
Грибы могут размножаться вегетативным, бесполым и половым путем.
Вегетативное размножение
осуществляется при помощи кусочков мицелия, плодовых тел, а также с помощью оидий и хламидоспор. Отдельные части мицелия, попадая на новый субстрат, разрастаются и дают начало новой особи.
Клетки некоторых грибов могут распадаться на отдельные округлые или овальные клетки - оидии. Такие клетки могут давать начало новому мицелию или почковаться.
В отличие от оидий хламидоспоры имеют толстую оболочку и большое количество питательных веществ, вследствие чего способны долгое время переносить неблагоприятные условия, не теряя жизнеспособности.
Бесполое и половое размножение осуществляется при помощи спор. К органам бесполого размножения
относятся зооспорангии с зооспорами, спорангии со спорангиоспорами и конидиеносцы с конидиями. При бесполом размножении споры образуются без оплодотворения.
Зооспорангии представляют собой вытянутые или округлые клетки, внутри которых в результате деления цитоплазмы и ядра образуются споры с одним или двумя жгутиками. Жгутики служат для перемещения в воде, а споры называются зооспорами.
Спорангии также представляют собой особые вздутия, расположенные на особых гифах - спорангиеносцах. Внутри них образуются одноклеточные бесцветные споры, называемые спорангиоспорами.
В отличие от перечисленных органов размножения конидии на конидиеносцах образуются экзогенно, т.е. снаружи. Конидиеносцы могут быть простыми или разветвленными и характерны для разных видов грибов. Конидии на них образуются поодиночке, цепочками или головками. В большинстве случаев конидиеносцы развиваются не отдельно, а образуют определенные структуры - коремии, ложа, пикниды.
При половом размножении
появлению споры предшествует слияние разнополых клеток. Наиболее простым вариантом полового размножения считается изогамия, при которой сливаются две одинаковые по размеру, но разные по полу зооспоры, в результате чего образуется циста.
Более сложные варианты полового размножения - оогамия и зигогамия. В первом случае на мицелии образуются мужские (антеридии) и женские (оогонии) половые органы. После слияния их содержимого образуется ооспора. Во втором случае происходит слияние двух клеток-гаметангиев, образовавшихся на разнополых гифах. В результате образуется бугристая зигоспора. Циста, ооспора и зигоспора характерны для низших грибов и являются покоящимися спорами, способными переносить неблагоприятные условия.
У высших грибов половой процесс представлен гаметангиогамией или соматогамией, завершающихся образованием соответственно сумок и базидий. Сумка - это округлая булавовидная или цилиндрическая клетка, внутри которой развиваются половые споры. Базидия - это булавовидная клетка, на выростах которой формируются базидиоспоры.
Существуют грибы, в жизненном цикле которых имеется только половое или только бесполое поколение, у других эти поколения чередуются. При этом, как правило, бесполое спороношение развивается в течение вегетационного сезона, а бесполые споры служат для расселения гриба. Появление полового спороношения свидетельствует об окончании вегетативного роста, изменении условий среды. Распространяются споры пассивно при помощи ветра, воды, насекомых, человека, а также вместе с растениями и их частями.
Как и все живое на Земле, гриб начинает свою жизнь с одноклеточного состояния — споры . Размеры спор грибов не превышают 10-25 мк в поперечнике.
Внешний вид их поразительно разнообразен. Достаточно сказать, что внешний вид спор служит одним из важных систематических признаков классификации грибов.
Спора прорастает, путем последовательного митотического деления разрастается в длину, образуя тонкие нити-гифы.
Гифы , вырастающие из одной споры, расходятся от нее во все стороны лучами, ветвятся и образуют первичный мицелий.
Дальнейшее развитие однополого первичного мицелия шляпочных базидиальных грибов включает своеобразную форму полового процесса. При соприкосновении кончиков гиф двух разнополых мицелиев происходит разрушение оболочек и слияние протопластов. Ядра сливающихся клеток не соединяются, и образуется двухъядерная клетка — дикарион. Этот процесс именуется плазмогамией . При дальнейшем делении двухъядерных клеток возникает своеобразный феномен, обеспечивающий каждой клетке получение и мужского и женского ядра. В начале деления на одном — заднем — конце клетки гифы образуется отросток — "крючок", или "пряжка", и переднее ядро спускается в этот отросток, после чего каждое из ядер делится на два. Одно из дочерних ядер переднего ядра из "крючка" отходит вперед, в переднюю клетку, где остается и одно из дочерних ядер заднего ядра после образования перегородки. При этом дочерние ядра заднего и переднего ядра вместе с "крючком" остаются в задней клетке. Так образуются две клетки, каждая с двумя разнополыми ядрами.
Из этих клеток путем последовательного деления разрастается вторичный двухъядерный мицелий. Он может при благоприятных внешних условиях образовать наземное плодовое тело - гриб. Плодовое тело представляет собой компактное сплетение гиф и служит органом для образования спор.
Гифы грибного мицелия могут, особенно у дереворазрушающих грибов, соединяться в толстые образования - шнуры-ризоморфы, которые в почве или на ином субстрате тянутся очень далеко. Иногда гифы образуют компактные темные образования — склероции. В этих и других разновидностях мицелия гифы никогда не сливаются, идут тесно и параллельно, как и в плодовом теле.
Споры у базидиальных шляпочных грибов образуются на нижней стороне шляпки. Здесь под мякотью шляпки вырастает гименофор (несущий гимений), состоящий из пластинок или трубочек, иногда просто складок, спускающихся на ножку, или шипиков и сосочков. На гименофоре развивается спорообразующий слой — гимений. В нем гифы заканчиваются и образуют концевые удлиненные двухъядерные клетки своеобразного строения — в форме цилиндра, булавы и др., — именуемые базидиями. Между плодоносящими базидиями располагаются тоже удлиненные клетки гиф - цистиды и парафизы. Эти одноклеточные или многоклеточные образования в форме мешочков бесплодны и выполняют, вероятно, питательную функцию в отношении базидий.
Процесс образования базидиоспор начинается со слияния обоих ядер базидий в одно большое, что трактуется как половой процесс — кариогамия. Путем быстрых последовательных делений большое ядро превращается в четыре дочерних, отделяющихся друг от друга перегородками. Далее каждое из новых ядер проникает в один из концевых отростков базидии — через стеригму, на конце которой происходит в течение нескольких часов созревание спор.
Плодовое тело, гименофор, гимений и базидии следует рассматривать как производные мицелия.
Описанный выше процесс размножения свойствен почти всем базидиальным грибам, но встречаются и отклонения от него. Так, у гриба Coprinus sp. первичный мицелий путем слияния своих же клеток становится двухъядерным, способным к развитию плодовых тел. У некоторых видов клетки мицелия могут содержать одно двуполое ядро, у других не образуется "пряжка". Образование "пряжки" характерно для размножения многих высших базидиальных грибов.
Для сравнения кратко упомянем о способах размножения других групп грибов. Споры образуются уже у слизевиков, которых раньше относили к классу низших грибов, а теперь выделяют в самостоятельный тип. Обычно слизевики существуют в виде одноядерных гаплоидных голых амеб, питающихся бактериями и способных к размножению. В определенный момент все амебы сливаются в один большой многоядерный плазмодий, в котором образуются споры, представляющие собой не что иное, как те же инцистированные амебы. При прорастании спор из них вытекают амебы. Цикл развития истинных слизевиков — миксомицетов, а также хитридиомицетов, оомицетов и, наконец, аскомицетов (сумчатых грибов) довольно сложен и включает в себя как фазу бесполого, так и фазу полового размножения. У многих из них образуются половые органы — оогонии и антеридии.
Грибы отличаются своим необычайно обильным плодоношением. Так, плодовое тело шампиньона - Agaricus campestris величиной в 8 см в поперечнике образует 1800 млн спор, выдавая каждую секунду по 40 млн спор. На 214 пластинках гриба Coprinus sp. образуется до 5000 млн спор, по 100 млн в час, а еще больше — до 1 000 млн спор имеется у ежовика — Sarcodon imbricatus.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter .
Особенность грибов в том, что любое их размножение (вегетативное, бесполое либо половое) обязательно связано с процессом образования спор, который характеризуется исключительно высокой плодовитостью. Количество спор, которое производят грибы, исчисляется миллиардами.
Такое размножение и расселение грибов с помощью спор, на первый взгляд, сближает их с семенным размножением растений. Но на самом деле здесь имеет место только кажущееся сходство, связанное с прикрепленным образом жизни и грибов, и растений.
У грибов широко распространено вегетативное размножение - с помощью нитей грибницы, или мицелия, которые могут распространяться самостоятельно, а так же механическим путем, токами воздуха, капельной влагой, различными животными и насекомыми.
Споры грибов, развивающиеся в период спороношения, могут отличаться. Одни споры - пропагативные - развиваются в огромном количестве, служат для быстрого расселения грибов. Так, в плодовом теле дождевика - до 7,5 миллионов спор, а шампиньон за 5 дней рассеивает более 10 биллионов спор. Такие споры быстро прорастают, но являются маложивучими, часто недозрелыми и в больших количествах погибают. Другие споры - покоящиеся - развиваются в малом количестве, нуждаются в покое, во время которого происходит их дозревание, служат для сохранения вида при неблагоприятных условиях. Такие споры могут сохраняться до 10-12 лет и прорастают медленно. Прорастают они обычно спорангием, дающим массу спор.
Вся окружающая среда - воздух, почва, вода - всегда переполнены спорами грибов. Спор много на растениях, животных. Вот почему грибы внезапно появляются там, где для них складываются благоприятные условия. Американские микологи в районе Великих озер проводили изучение условий распространения спор ржавчинного гриба - возбудителя опасного заболевания сосны - пузырчатой ржавчины. Было определено, что споры гриба выделялись круглосуточно. Споры, освобождающиеся ночью, меньше и легче спор, освобождающихся днем, и поэтому лучше распространялись токами воздуха в ночных условиях. В атмосфере, таким образом, путешествовали тучи спор, довольно устойчивых и обнаруживающихся в основном ночью. Высокий поток воздуха нес эти споры на 16-27 километров в сторону от Великих озер, там споры оседали и вызывали массовое заражение деревьев сосны. Часто грибные споры разносятся зверями, человеком, насекомыми.
Интересными являются некоторые сумчатые грибы, развивающиеся на навозе теплокровных животных - Аскоболус, Пилоболус, Копринусы. Например, споры Пилоболуса не могут прорасти, если не прошли через кишечник теплокровного животного, едкий желудочный сок и температура 37°С способствуют их прорастанию. Развиваясь на навозе, грибок образует массу спорангиев, сидящих на тоненькой ножке. При созревании спорангий вздувается, с силой отскакивает вверх и прочно прилипает к травянистому растению, с которого попадает в кишечник животного. У гриба Аскоболуса споры имеют слизистые придатки, которые крепко прикрепляют спору к травянистому стеблю. Некоторые навозники выделывают огромное количество спор. Так, навозник Копринус косматус за час образует до 100 миллионов спор. Его жизнь коротка (всего 48 часов), и за это время нужно дать жизнь своему потомству.
Известно, что птицы являются переносчиками многих болезней растений. Они занесли из Америки в Европу патогенный гриб возбудителя американской мучнистой росы крыжовника. Птицы разносят в лесу болезнь шютте на сеянцах сосны и кедра, эксобазидиум - на бруснике, распространяют также многие мучнисто-росяные и ржавчинные грибы. Птицы, особенно из семейства врановых, любят поедать грибы.
В расселении грибов большую роль играют звери, живущие в лесу. Многие дикие звери - большие любители грибов. Отлично разбираются в грибах белка и бурундук. Белка собирает подосиновики, подберезовики, ежевики. С любовью накалывает их на веточки, сучья, сушит и складывает в свое дупло. При этом рассеиваются их споры. Сборы белок значительны, достигают иногда до килограмма отличных сушеных грибов. Мышевидные грызуны любят сыроежки, рыжики, подберезовики, но пренебрегают белыми и подосиновиками. Нередко мыши являются распространителями домового гриба, этого злостного врага деревянных построек. Хорошими помощниками в распространении спор грибов являются насекомые и черви. Разрыхляя почву, земляные черви все время разносят споры грибов. Им помогают многочисленные личинки насекомых, которые находятся в почве или поселяются в плодовых телах грибов. Питаясь мякотью гриба личинки грибных комариков распространяют его споры. Поэтому червивость плодового тела полезна для жизни гриба.
Большую роль в распространении спор грибов играет, конечно же, и ветер. Он разносит споры дождевиков, многих шляпочных, ржавчинных и головневых грибов. Воздушные течения разносят споры на расстояния до 1000 и более километров. Самой незначительной разницы температуры на почве и в кроне деревьев и кустарников достаточно для распространения спор грибов в слоях неравномерно прогретого воздуха.
Вот почему говорят: "Неподвижные грибы всегда в движении". Грибы появляются внезапно там, где складываются для них благоприятные условия, переселяются и путешествуют повсюду.
Опытный грибник бережно относится к этим бесценным дарам леса. Старый спороносящий гриб - семенник - он не бросит на землю, где гриб бесполезно сгниет, а наколет, повыше, на сук дерева или ветку кустарника, чтобы высохшие споры подхватил ветер и разнес по лесу.
При использовании материалов сайта, необходимо ставить активные ссылки на этот сайт, видимые для пользователей и поисковых роботов.
У грибов различают два типа размножения: вегетативное и репродуктивное.
Вегетативное размножение осуществляется частями вегетативного тела или спорами, которые образуются клетками вегетативных гиф (рис. 16). Самая примитивная форма вегетативного размножения - частицами гиф (а также обрывками шнуров, ризоморф, склероциями), которые, будучи отделены от материнского мицелия и попав в благоприятную среду, могут дать начало новому самостоятельному мицелию.
У некоторых грибов, например дрожжевых, вегетативное размножение осуществляется почкованием клеток. На определенном этапе развития на них появляются небольшие выросты, которые постепенно увеличиваются в размерах, затем отделяются от материнской клетки и в свою очередь начинают почковаться. Такие клетки называют бластоспорами.
Своеобразная форма вегетативного размножения - образование оидий и хламидоспор. Оидии образуются в результате распадения гиф, начиная с их концов, на отдельные членики, которые в дальнейшем дают начало новому мицелию. Оидии снабжены тонкой оболочкой и недолговечны. Они встречаются у голосумчатых и других грибов. Разновидностью оидий являются геммы, отличающиеся от них более плотной темной оболочкой и способностью дольше сохраняться (например, в зимний период). Геммы известны у многих сумчатых, несовершенных, а также некоторых головневых грибов.
Хламидоспоры возникают путем уплотнения и обособления содержимого отдельных клеток мицелия, которое при этом покрывается толстой темноокрашенной оболочкой. Освободившиеся из клеток материнских гиф хламидоспоры способны долго сохраняться при неблагоприятных условиях. Прорастая, они образуют органы спороношения или мицелий. Хламидоспоры образуются у многих базидиомицетов (например, головневых), дейтеромицетов, оомицетов.
Репродуктивное размножение происходит с помощью спор, которые образуются внутри или на поверхности специальных органов, отличающихся по своему строению от вегетативных гиф. Репродуктивное размножение может быть бесполым - с образованием спор без оплодотворения, и половым, при котором образованию спор предшествует половой процесс.
Бесполое размножение. Как правило, бесполое спороношение грибов формируется на хорошо развитом мицелии, имеющем достаточный для образования спор запас питательных веществ. Спороносными органами при бесполом размножении грибов служат специальные ответвления или участки гиф, внутри или снаружи которых образуются споры (рис. 17).

Простейшая форма бесполого размножения низших грибов - образование зооспорангиев, представляющих собой расширенные окончания гиф. Их содержимое распадается на подвижные споры с одним или двумя жгутиками - зооспоры, которые могут передвигаться и сохранять жизнеспособность только в воде. Поэтому зооспоры свойственны водным и некоторым почвенным грибам, а у наземных видов, сохранивших связь с водной средой, образуются во время дождя, росы или густого тумана.
Другой формой бесполого размножения низших грибов являются спорангии - шаровидные вместилища на концах ответвлений мицелия. Ветвь, несущая спорангий, называется спорангиеносцем. Внутри спорангия формируются неподвижные одноклеточные споры - спорангиоспоры. Они одеты твердой оболочкой; образуются у мукоровых грибов.
Наиболее распространенная форма бесполого размножения, свойственная почти всем высшим и некоторым низшим грибам,- конидиальное спороношение. Конидиями называют споры, образующиеся на конечных ответвлениях специальных органов - конидиеносцев. Конидиеносцы и конидии очень разнообразны по форме, размерам, строению и окраске, а также по характеру их развития и размещения. Разновидностями конидиального спороношения грибов являются коремии, ложа и пикниды.
Коремии - это пучки плотно соединенных друг с другом конидиеносцев, на концах которых образуются конидии. Скопления конидий на коремиях могут быть порошащими или слизистыми. Спороношения коремального типа часто встречаются у грибов из порядка гифомицетов, у сумчатых грибов.
Ложа представляют собой подушковидные, более или менее плотные мицелиальные образования, формирующиеся на поверхности пораженного субстрата или погруженные в него, но при созревании выступающие наружу. Конидиеносцы располагаются на поверхности ложа сплошным слоем. Такая форма конидиального спороношения характерна для грибов порядка меланкониевых.
Пикниды - это шаровидные (иногда немного приплюснутые) или грушевидные вместилища с плотной, обычно темной оболочкой. Они бывают одиночными (иногда их называют однокамерными) или образуются группами в общей строме (многокамерные пикниды). Конидии в пикнидах, или пикноспоры, образуются на коротких конидиеносцах, покрывающих внутреннюю стенку пикниды. Созревая, они выделяются из пикниды через специальное отверстие (устьице) сплошной слизистой массой, застывающей на воздухе в виде капель или скрученных нитей, нередко имеющих яркую окраску. Чаще всего пикниды частично или полностью погружены в питающий субстрат и наружу выступают лишь их верхние части или устьица. Спороношение типа пикнид - основной признак грибов порядка пикнидиальных.
У многих фитопатогенных грибов в течение вегетационного периода наблюдается несколько генераций бесполого спороношения; этим обеспечиваются массовое распространение инфекционного начала и повторное заражение растений.
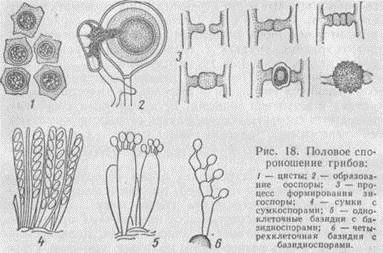
Половое размножение. Половой процесс у грибов весьма разнообразен (рис. 18). В простейшей форме он представлен изогамией, т. е. слиянием двух одинаковых по форме, но разнополых зооспор. В результате формируется циста. У некоторых наиболее примитивных грибов (например, слизевиков) продуктом полового процесса типа изогамии является диплоидный амебоид.
Более сложные формы полового размножения - оогамия и зигогамия. При оогамии на мицелии закладываются различные по форме и строению половые клетки: женская - оогоний и мужская - антеридий. После слияния их содержимого образуется ооспора. При зигогамии сливается содержимое двух внешне одинаковых клеток разнополых мицелиев. В результате образуется зигоспора. Цисты, ооспоры и зигоспоры - это покоящиеся споры, покрытые толстой оболочкой и предназначенные для сохранения вида при неблагоприятных условиях. Они характерны для низших грибов.
У высших грибов (сумчатых и базидиальных) половое воспроизведение завершается формированием сумок или базидий. Сумки (аски) - это различной формы мешковидные клетки, внутри которых развиваются сумкоспоры (аскоспоры). Обычно сумка содержит восемь сумкоспор. Базидия представляет собой тело булавовидной или цилиндрической формы, на поверхности которого образуются базидиоспоры. Чаще всего их четыре.
Грибы, у которых мицелий наделен свойствами обоих полов и половой процесс осуществляется путем слияния цитоплазмы и ядер клеток одного и того же мицелия, называются гомоталличными. Грибы, у которых каждый мицелий обладает свойствами только одного пола, а в половом процессе участвуют клетки и ядра разнополых мицелиев, называются гетероталличными. В связи с тем что мицелии разного пола у гетероталличных грибов морфологически неразличимы (у многих видов нет четких морфологических различий даже между копулирующими клетками), конкретный пол у грибов часто не указывают и половую принадлежность отдельных особей обозначают знаками плюс (+) и минус (-).
Переход к половому размножению у грибов часто бывает связан с окончанием вегетативного роста, наступлением неблагоприятных условий, перезимовкой. У фитопатогенных видов половое спороношение, как правило, наблюдается один раз в течение года, а споры, образующиеся половым путем, осуществляют первичное заражение растений весной или в начале лета.
Способность грибов формировать в цикле развития разные типы спороношений называется плеоморфизмом. Циклы развития большинства грибов включают два спороношения - половое и бесполое, которые чаще всего имеют самостоятельные видовые названия. В то же время существует немало видов, имеющих наряду с половым несколько различных бесполых спороношений. Такой цикл развития называют сложным. Примером грибов со сложным циклом развития могут служить ржавчинные грибы. Дейтеромицеты (несовершенные грибы) имеют только бесполые спороношения. У некоторых сумчатых и базидиальных грибов известно лишь половое спороношение.
Споры грибов, предназначенные для распространения и быстрого прорастания в течение вегетационного периода, называются пропагативными. Такие споры имеют тонкие, обычно бесцветные оболочки и не могут долго сохранять жизнеспособность. Споры, служащие для длительного сохранения при неблагоприятных условиях, называются покоящимися. Они бывают снабжены толстой и, как правило, пигментированной оболочкой.
Спорообразующая способность большинства грибов очень велика. Например, у мукоровых грибов один спорангий может содержать несколько десятков тысяч спорангиоспор. В каждой пустуле ржавчинных грибов созревает несколько сотен спор. Число сумкоспор в плодовом теле некоторых сумчатых грибов исчисляется миллионами, а плодовые тела многих трутовых грибов выделяют миллиарды базидиоспор ежедневно в течение всего периода споруляции. Количество продуцируемых грибами спор многократно увеличивается соответственно числу генераций бесполого спороношения.